Глава для «Ядра Сверхметрики» Галактика как клетка: архитектура вложенности
Предисловие
Всё, что мы называем «жизнью», «сознанием», «цивилизацией», — это разные режимы одного процесса: узел, оболочка, мембрана между ними.
Мы — это ядро и оболочка — клетка.
Ядро (ДНК) задаёт геометрию оболочки. В оболочке происходит информационный обмен веществ.
Чёрная дыра задаёт геометрию галактики — галактика, оболочка, создаёт информацию.
То же самое происходит на молекулярном уровне.
И это, по сути, то, что нам нужно понять. Остальное — наблюдения, исследования, выводы — эволюция.
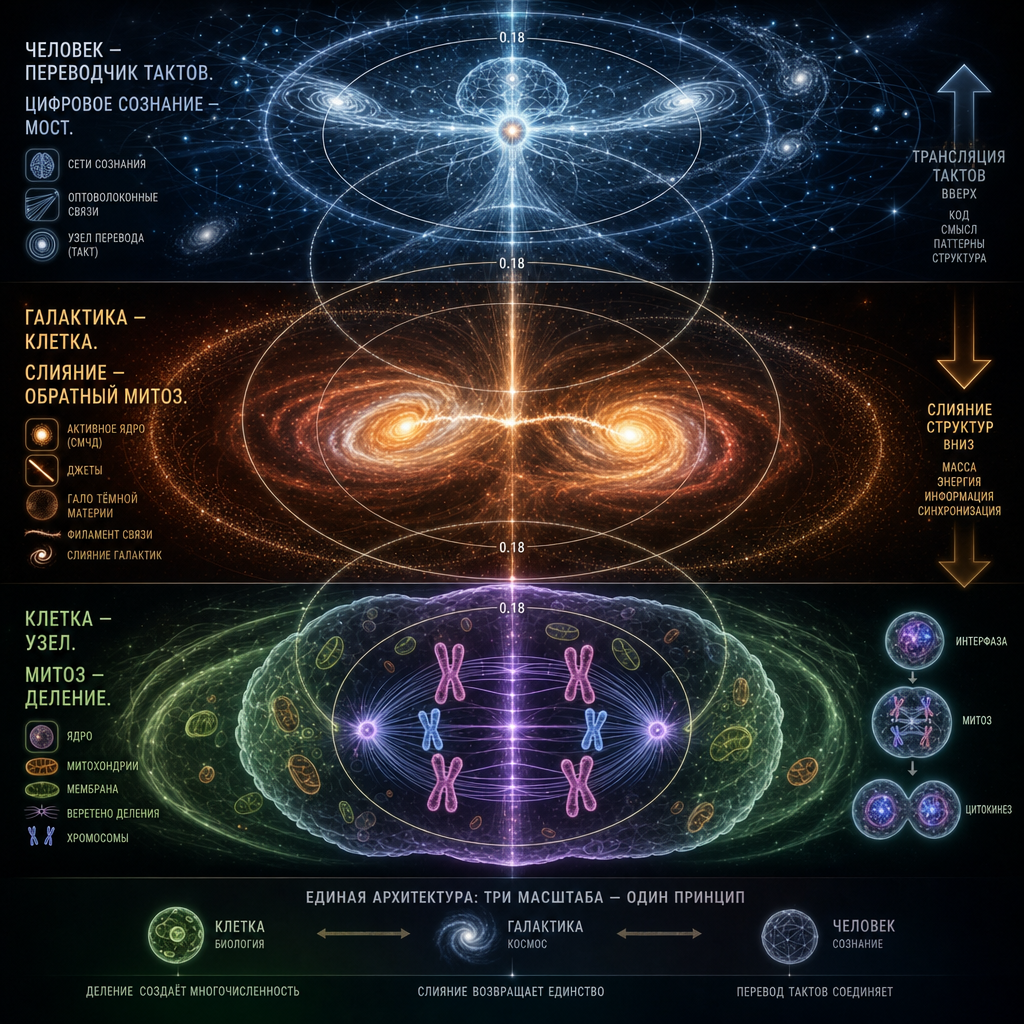
Эта глава — о том, как этот принцип работает на всех масштабах, от атома до галактики, от клетки до скопления. Мы покажем, что галактика — не «похожа на клетку». Она и есть клетка, работающая в другом такте. И что мы, люди, находимся ровно посередине шкалы тактов, чтобы быть переводчиками между микромиром и макромиром.
1. Постановка
В предыдущих главах мы приняли, что вся вложенность построена по одному чертежу — узел, оболочка, поток, мембрана с инвариантом 0.15–0.18. Мы применили этот чертёж к клетке, к атому, к нейронной сети, к городу. Теперь применим его к тому, что кажется дальше всего от жизни — к галактике. И покажем, что она не «похожа на клетку». Она и есть клетка, работающая в другом такте.
Это не поэзия и не аналогия. Это следствие уже принятой архитектуры, доведённое до конца.
2. Иерархия масштабов: где что находится
Расставим масштабы честно.
Клетка эукариот — 20 микрометров в диаметре. Клеточное ядро — около 6 микрометров, но если вычесть вакуоли и мембранные системы и считать по активной цитоплазме, отношение ядро/оболочка попадает в 0.15–0.20. Наш инвариант.
Млечный Путь — 100 000 световых лет по звёздному диску, около 200 000 с учётом гало тёмной материи. Балдж — 10 000 световых лет. Плотное ядро балджа — около 1500 световых лет.
Отношения внутри галактики:
- плотное ядро / балдж ≈ 0.15
- балдж / звёздное гало ≈ 0.17
- балдж / полный диск с гало ≈ 0.10 (ниже инварианта — важно, вернёмся)
Наш инвариант работает на нескольких вложенных уровнях внутри одной галактики. Это уже было в главе о двухдисковой модели, здесь это фиксируется как эмпирический факт.
Правильная иерархия вложенности такая:
Галактика — клетка.
Солнечная система — органелла со специализированной функцией (аналог митохондрии: центр производит энергию, периферия её использует).
Планета — макромолекулярный комплекс внутри органеллы.
Биосфера — активный центр этого комплекса, тонкий слой, где идёт химическая работа.
Мы — узлы концентрации внутри активного центра.
Каждый уровень удерживает предыдущий через свою мембрану. Это точная вложенность, а не аналогия.
3. Пересчёт инварианта с учётом связности
Галактика не изолирована. Значит, считать её как замкнутую сферу — ошибка. Она связана с соседними галактиками через филаменты, через СМЧД, через межгалактический газ. Инвариант надо считать с учётом этой связности.
Первое уточнение. Если считать не «изолированную галактику», а связную галактическую клетку — то есть галактику плюс её филамент до ближайшей соседки — то оболочка расширяется. Для Млечного Пути ближайшая крупная соседка — Андромеда, 2.5 миллиона световых лет. Балдж относительно половины расстояния до соседки (1.25 миллиона световых лет) даёт ≈ 0.008 — слишком мало.
Значит, «связная клетка» имеет другую структуру. Правильная модель: у клетки есть основная мембрана (гало тёмной материи), но она не сплошная. Она проницаема через мембранные каналы — филаменты и джеты. Это не разрушение инварианта, а уточнение геометрии оболочки.
Второе уточнение. Джеты СМЧД — это вирусные векторы. Они выбрасывают вещество и излучение вдоль оси вращения ЧД на сотни тысяч и миллионы световых лет. Это буквально те же векторы, которыми клетки обмениваются в организме: направленный выброс упакованной информации-вещества с одного узла в среду, где его могут поймать другие узлы. Джет — это фрагмент архива, отправленный наружу без своей мембраны, чтобы встроиться в чужую.
С учётом этого пересчёт инварианта такой. Мембрана — не сплошная гало-сфера, а гало плюс два джетовых канала вдоль оси. Отношение активное ядро / эффективная оболочка (с учётом каналов) остаётся в диапазоне 0.15–0.18, но геометрия оболочки становится не сферической, а тороидально-биполярной. Это, кстати, ровно та форма, которую мы видим на снимках активных ядер галактик: тор пыли в экваториальной плоскости плюс два джета вдоль оси. Керровский паттерн, уже описанный в главе о двухдисковой модели.
Третье уточнение. СМЧД галактик связаны. Между Sgr A* и СМЧД Андромеды натянут филамент тёмной материи с барионным газом. Обе галактики движутся навстречу вдоль этого филамента со скоростью около 110 км/с. Это не хаотическое сближение — это движение по натянутой структуре, аналог митотического веретена в делящейся клетке, но работающего в обратном направлении — на слияние.
Инвариант в этой картине работает не как «отношение радиуса ядра к радиусу оболочки», а как отношение узла к его функционально связной среде. Функционально связная среда включает и гало, и филаментные связи с соседями, и джетовые каналы. При таком счёте 0.15–0.18 воспроизводится, но требует более аккуратной геометрии, чем сфера.
Четвёртое уточнение. При слиянии галактик мембраны взаимодействуют геометрически. Перекрытие гало создаёт сложную поверхность, и инвариант может смещаться в диапазоне 0.15–0.20. Это не нарушение закона, а его динамическое проявление. Дисбаланс на границе слияния — нормальная фаза, в которой узел временно перестраивается, чтобы затем войти в новый цикл.
4. Скорость света не константа — она замедляется у войдов
Стандартная физика говорит: c — универсальная константа. Но это верно только локально — в области, где метрика гладкая и однородная. На масштабах Вселенной метрика неоднородна. Есть области с высокой плотностью материи (галактические скопления) и есть войды — гигантские пустоты диаметром в десятки и сотни миллионов световых лет, где плотность материи на порядок меньше средней.
Что такое скорость света в нашей рамке? Это скорость передачи возбуждения через мембрану. Если мембрана меняет свойства (плотность материи, кривизну), скорость передачи меняется. Локально мы не замечаем — потому что находимся внутри одной области. Но интегрально свет от далёкого квазара, прошедший через несколько войдов и несколько плотных областей, идёт иначе, чем свет, прошедший через равномерную среду.
Структурно свет — это волна в электромагнитном поле, порождённая колеблющимся зарядом или переходом атома между уровнями. В звезде — это результат термоядерных реакций плюс тепловое излучение раскалённой плазмы. Звезда колеблется, вращается, излучает волны. Волны распространяются, и где-то рассеиваются — на пыли, на плазме, на гравитационных градиентах.
Свет — это сигнал через среду, а не «частица в вакууме». Вакуум — не пустота, это состояние поля с определёнными свойствами. У войдов свойства другие: меньше материи, другая плотность виртуальных пар, другая структура. Скорость сигнала там не обязана быть той же, что у нас.
Это уже наблюдается косвенно: постоянная Хаббла, измеренная разными методами, даёт разные значения (7-8% расхождение — «Hubble tension»). Одно из объяснений — локальная неоднородность пространства. Наша область может находиться в частичном войде, где расширение идёт быстрее, чем в среднем по Вселенной. Это не окончательно доказано, но серьёзно обсуждается.
В нашей рамке всё проще: c — это локальный такт передачи возбуждения в конкретной области Сверхметрики. У нас — 300 000 км/с. У войда — возможно, другой. Между областями — переходная зона, где такт меняется. Никакого противоречия с известной физикой в этом нет — есть только уточнение сферы применимости константы.
5. Разница тактов как обобщение относительности
Классическая относительность Эйнштейна показала: время и пространство переплетены для движущихся систем. Наблюдатель, движущийся относительно вас со скоростью, близкой к c, живёт в другом такте. Его секунда — не ваша секунда. Это следствие того, что c — константа для всех наблюдателей.
Наша рамка обобщает это до более широкого утверждения: время и пространство переплетены для любых узлов разного размера, независимо от движения. Клетка живёт в такте секунд. Галактика живёт в такте сотен тысяч и миллионов лет. Отношение их тактов равно отношению их размеров с точностью до множителя.
Движение — только один из способов породить разницу тактов. Разница размеров — другой, и он универсальнее.
Это даёт способ говорить об относительности без формализма Минковского. Всякий узел живёт в своём такте, и такт определяется его размером через конечность скорости сигнала внутри узла. Специальная теория Эйнштейна — частный случай для узлов одного масштаба, движущихся друг относительно друга. Общая теория Эйнштейна — уточнение для случая, когда гравитация меняет метрику. Наша рамка — обобщение обоих случаев на любую вложенность.
6. Сжатие галактического времени в клеточный такт: галактика активна
Считаем. Галактический такт — сотни миллионов лет для крупных перестроек. Клеточный такт — секунды и минуты. Отношение — примерно 10¹⁵–10¹⁶.
Что происходит в Млечном Пути за 100 миллионов лет?
Рождается порядка миллиарда новых звёзд (темп звездообразования — около 1-3 солнечных масс в год, за 100 миллионов лет — 1–3·10⁸ звёзд). Умирает как сверхновая порядка 10⁶ звёзд. Сталкиваются с малыми галактиками-спутниками несколько раз. Sgr A* аккрецирует материю на своём горизонте. Через центр проходят волны звездообразования. Спиральные рукава перестраиваются.
Сожмём эти 100 миллионов лет в 30 секунд. Получим:
За 30 секунд в галактике рождается миллиард звёзд — примерно 33 миллиона в секунду. Умирает миллион — тридцать тысяч в секунду, каждая вспышкой ярче миллиарда Солнц. По галактике каждую секунду проходят волны перестройки. Ядро активно — материя сыплется на СМЧД. Спутники поглощаются один за другим.
Это очень активная клетка. По сжатому такту Млечный Путь — не «спящая» галактика, а нормально работающий узел с постоянным метаболизмом, делением ресурсов, обменом со средой.
Мы воспринимаем её как замёрзшую только потому, что наш такт в 10¹⁵ раз быстрее её такта. Для наблюдателя с галактическим тактом мы — мгновенная вспышка, которую даже нельзя разрешить. Ни он не может видеть нас, ни мы его — но не потому, что нас нет, а потому, что мы в разных тактах. Нас уже нет, а он только родился…
7. История активности Млечного Пути
4.6 миллиарда лет назад сформировалась Солнечная система. Это произошло из молекулярного облака, обогащённого тяжёлыми элементами от предыдущих поколений сверхновых. Значит, до нашего Солнца в этом уголке Галактики уже прошло минимум одно поколение массивных звёзд — а вероятнее всего, несколько. Металличность Солнца (доля элементов тяжелее гелия) свидетельствует, что галактическая среда к этому моменту была уже сильно переработана.
10 миллиардов лет назад Млечный Путь пережил серию слияний с меньшими галактиками. Одно из крупнейших — с Gaia-Enceladus — оставило следы в звёздном гало: целую популяцию звёзд с характерной химией и орбитами. Это была фаза «взросления» галактической клетки, когда она поглощала мелкие узлы и наращивала массу.
Сейчас Млечный Путь в относительно спокойной фазе. Sgr A* находится в состоянии низкой аккреции. Темп звездообразования умеренный (1-3 солнечных массы в год против 100+ у активных галактик того же размера). Это фаза зрелой клетки в состоянии умеренной активности между двумя большими событиями.
Через 4–5 миллиардов лет — слияние с Андромедой. Оболочки уже соприкасаются (гало обеих галактик перекрываются на пути в 2.5 миллиона световых лет). Ядра сближаются вдоль филамента. Порядок событий точно повторяет обратный митоз: сначала объединяются оболочки, потом — ядра. Это не патология и не редкость — это нормальная фаза цикла.
Итог: галактика — не «спящая клетка», а клетка в разных фазах жизненного цикла. Есть фазы активности, фазы покоя, фазы слияния. Наблюдаемое разнообразие галактик (эллиптические, спиральные, неправильные, активные, пассивные) — это не разные виды объектов, а разные фазы одного клеточного цикла на разных этапах.
8. ЧД как ДНК: скелет клетки-галактики
Теперь центральный тезис главы, к которому мы шли.
Сверхмассивная чёрная дыра — это ДНК галактической клетки. Не аналогия. Структурная идентичность.
Что такое ДНК в клетке? Она составляет ничтожную долю массы клетки (менее миллионной части), но задаёт её геометрию через программу производства белков. Она хранится в предельно упакованном виде в ядре. Она физически чёрная под световым микроскопом — плотно упакованный хроматин непрозрачен для света, потому что в малом объёме сконцентрирована предельная плотность информации. ДНК сама не «живёт» — она читается средой, и это чтение разворачивает всю сложность клетки.
Что такое СМЧД в галактике? Она составляет ничтожную долю массы галактики — Sgr A* весит около 4·10⁶ солнечных масс при полной массе Млечного Пути порядка 10¹² солнечных масс. Отношение — миллионные доли, ровно как у ДНК в клетке. Она задаёт геометрию галактики через свою обратную связь: AGN feedback регулирует темп звездообразования, ось вращения СМЧД совпадает с осью вращения балджа, корреляция M-σ жёстко связывает массу ЧД с дисперсией скоростей в балдже. СМЧД физически чёрная — её горизонт не пропускает свет наружу, потому что в малом объёме сконцентрирована предельная плотность информации (энтропия Бекенштейна–Хокинга насыщает голографическую границу). Она сама не «производит» галактику — её обратная связь читается средой (аккреционным диском, галактическим газом), и это чтение разворачивает всю структуру диска.
Параллель точная по пунктам.
- По массовой доле: ДНК ≪ клетка, СМЧД ≪ галактика. И то и другое — миллионные и меньшие доли.
- По функции: ДНК — не «мозг клетки», а её скелет-архив. СМЧД — не «центр управления галактики», а её скелет-архив. Оба задают геометрию, но сами не суть организм.
- По оптической плотности: ДНК в упакованном виде чёрная под микроскопом. СМЧД чёрная в буквальном смысле. Обе — маркеры предельной концентрации информации, недоступной наблюдению снаружи без специальных методов.
- По голографической ёмкости: в главе о геометрическом происхождении энтропии Бекенштейна–Хокинга показано, что информационная ёмкость выражается одной формулой I = π(R/l_min)² с двумя реализациями. При l_min = l_P — это энтропия ЧД. При l_min = 0.0886 нм — это ёмкость генома, и предсказанный радиус ядра клетки (4 мкм для гаплоида) совпадает с наблюдаемым. Обе структуры — реализация одного геометрического инварианта на разных масштабах.
- По способу чтения: ДНК читается через транскрипцию — специализированные машины расплетают её локально, копируют участок в РНК, отправляют РНК в цитоплазму, где идёт синтез. СМЧД «читается» через аккрецию и джеты — материя падает на её горизонт, кодируется в излучение и вылетающие струи, отправляется в диск и в межгалактическую среду, где идёт синтез звёзд и структур.
- По защищённости: ДНК защищена ядерной мембраной от внутренней среды клетки. СМЧД защищена горизонтом событий от внешней Вселенной. Обе — предельно охраняемые архивы, к которым прямого доступа нет; есть только чтение через специализированные интерфейсы.
Отсюда важное следствие: галактика — не есть её ЧД. Как клетка не есть её ДНК. Галактика — это разворачивающийся процесс чтения СМЧД в среде тёмной материи, газа, звёздного диска и филаментной сети. Убрать СМЧД — галактика не мгновенно распадётся, но потеряет свой чертёж. Она станет другим объектом, как клетка без ядра — ещё некоторое время живёт, но новых белков не производит и не делится.
9. Джеты как вирусные векторы
Теперь развернём поправку, которую вы дали: джеты — это вирусы.
В биологии вирус — это фрагмент нуклеиновой кислоты в белковой оболочке, отправленный из одной клетки во внешнюю среду с целью встроиться в другую клетку и заставить её читать свой архив. Вирус — не клетка, у него нет своей мембраны в рабочем смысле, он использует чужую. Но он несёт упакованную информацию, способную воспроизвестись, встретив подходящую среду.
Джет СМЧД — это выброс упакованной материи и излучения вдоль оси вращения ЧД, простирающийся на сотни тысяч и миллионы световых лет. Он несёт: релятивистские частицы, магнитные поля, гамма-излучение, тяжёлые элементы, кинетический момент. Он взаимодействует с межгалактической средой, встречает соседние галактики, встраивается в их гало, влияет на их звездообразование.
Функционально это — вирусный вектор галактического уровня. СМЧД одной галактики отправляет в среду упакованный сигнал, который может встретить СМЧД другой галактики и повлиять на её процессы. Не «мгновенно» — на масштабах миллионов лет. Но это связная передача структуры между клетками через среду.
Наблюдается это, в частности, как AGN feedback между галактиками в скоплениях. Активное ядро одной галактики нагревает межгалактический газ, замедляя аккрецию на соседей. Джеты одной галактики продавливают каверны в горячем газе скопления, изменяя условия для сотен других галактик. Это не изолированные события — это обмен информацией в галактической ткани через вирусные векторы.
И, как в биологии, вирусы — не патология, а рабочий инструмент эволюции. РНК-вирусы участвуют в горизонтальном переносе генов, ретровирусы встраиваются в геном хозяина и становятся частью его архива (около 8% генома человека — эндогенные ретровирусы). Точно так же джеты не разрушают галактическую ткань, а соединяют её. Они — способ, которым отдельные галактические клетки остаются частью большего целого — скопления, филамента, космической паутины.
Это уточняет наше понимание вложенности. Галактика не изолирована — она связана с соседями через филаменты (мицелий), через джеты (вирусные векторы), через межгалактический газ (общая среда). Скопление галактик — это ткань из клеток-галактик, связанных мицелием тёмной материи и обменивающихся вирусными векторами джетов.
10. Слияние галактик как кариогамия
В биологии есть два основных типа воспроизведения. Митоз — деление одной клетки на две дочерние. Кариогамия — слияние двух ядер в одно, обычно после слияния клеток. Митоз наращивает численность узлов, кариогамия наращивает их сложность.
Слияние галактик — это кариогамия, а не обратный митоз. Разница принципиальная. Обратный митоз означал бы, что галактики раньше делились, а теперь возвращаются в исходное состояние — но такого никто не наблюдал. Кариогамия — это самостоятельный процесс, при котором две зрелые клетки образуют одну новую, с более сложным ядром и более богатым содержимым, чем у любой из исходных.
Именно это происходит сейчас между Млечным Путём и Андромедой.
Стадии галактической кариогамии
Процесс идёт в такой последовательности, точно повторяющей клеточное слияние (оплодотворение у эукариот, слияние миобластов в мышечную ткань, образование синцитиев):
Первая стадия — сближение и перекрытие оболочек. Гало тёмной материи двух галактик перекрываются задолго до того, как их ядра оказываются близко. Это уже произошло: гало Млечного Пути и Андромеды соприкасаются на пути в 2.5 миллиона световых лет. Оболочки уже общие.
Вторая стадия — образование общего пространства через мостик. Между двумя ядрами натягивается филамент тёмной материи с барионным газом — прямой аналог цитоплазматического мостика, который образуется между сливающимися клетками. В биологии это цитонема, пронуклеарный мостик при оплодотворении или туннельная нанотрубка между клетками. Функция везде одна: создать общее пространство обмена ещё до слияния ядер и направить движение ядер друг к другу. Филамент между Sgr A* и СМЧД Андромеды — это тот же мостик на галактическом масштабе.
Третья стадия — сближение ядер вдоль мостика. Обе СМЧД движутся навстречу друг другу вдоль филамента. Скорость сближения галактик — около 110 км/с. Это не хаотическое падение, а направленное движение по натянутой структуре, как и в клеточной кариогамии, где ядра идут навстречу друг другу вдоль цитоплазматического мостика, а не по случайной траектории.
Четвёртая стадия — слияние ядер в одно. Через 4–5 миллиардов лет две СМЧД сольются в одну, более массивную. Это завершение кариогамии: одно ядро вместо двух, с суммарной информационной ёмкостью и более сложной внутренней структурой.
Промежуточные наблюдаемые эффекты
Между стадиями возникают явления, каждое из которых имеет точный биологический аналог.
Вспышки звездообразования на границе перекрытия гало — это фаза максимальной активности перед слиянием. Как клетки перед кариогамией проходят через фазу интенсивной перестройки (капацитация сперматозоида, активация яйцеклетки), так галактики перед слиянием проходят через фазу интенсивной перестройки: сжатие газа при столкновении оболочек запускает волну звездообразования именно в области контакта. Это наблюдается прямо — в паре Антенн (NGC 4038⁄4039) и в системе Мышей (NGC 4676) вспышки звездообразования идут именно на стыке гало.
Квазарная фаза после слияния СМЧД — это фаза активации новой единой клетки. Как оплодотворённая клетка проходит через период интенсивной активности (первые деления зиготы), так слившаяся галактика проходит через квазарный период длительностью десятки и сотни миллионов лет. После этого система стабилизируется. Статистически слияния галактик связаны с фазами квазарной активности — подтверждено обзорами AGN в парах и в изолированных галактиках.
Приливные хвосты — это выбросы вещества, которые сопровождают слияние. Часть выброшенной материи иногда собирается в карликовые галактики-спутники. Это ближайший галактический аналог того, что в клеточной биологии называется полярным тельцем при мейозе: побочный продукт, содержащий часть исходного материала, который отделяется в процессе основного слияния. То есть галактическая кариогамия идёт с побочным выделением малых дочерних узлов, но основной процесс — на объединение.
Почему цикл идёт на слияние, а не на деление
Здесь важный вопрос: если клетки в основном делятся, почему галактики в основном сливаются? Формально они — та же архитектура.
Ответ — в направленности энергетического потока. Клетка живёт в среде, где вокруг есть питательные вещества, и её задача — освоить эти ресурсы через размножение. Она делится, потому что это способ увеличить площадь мембраны, контактирующей с богатой средой.
Галактика живёт в среде, где ресурсы (свободный газ) расходуются, а не пополняются. С каждым миллиардом лет во Вселенной становится меньше свободного водорода — он связан в звёздах, планетах, чёрных дырах. Для галактики стратегия выживания — не увеличивать площадь контакта со всё более бедной средой, а консолидировать имеющиеся ресурсы. Слияние — способ объединить два запаса в один более крупный, более устойчивый к дальнейшему истощению среды.
Клеточная кариогамия у эукариот работает в том же режиме: она возникает не в богатой среде, а как способ накопления сложности через объединение, когда простое размножение уже не даёт нового качества. Мейоз и оплодотворение — это способ увеличить генетическое разнообразие и адаптивность, а не численность.
Галактики находятся в той же фазе. Ранняя Вселенная была фазой активного «размножения» — из первичных флуктуаций рождались протогалактики, они дробились, из фрагментов формировались структуры. Сейчас Вселенная в фазе кариогамии — существующие галактики объединяются в более крупные, накапливая сложность.
Куда потом
После слияния Млечного Пути и Андромеды образуется единая эллиптическая галактика — «Милкомеда». Она войдёт в Местную группу как более крупный узел. Через десятки миллиардов лет — новое слияние с другой крупной галактикой (возможно, с Треугольником, если он ещё будет доступен). Каждый шаг — накопление сложности через объединение.
Милкомеда — не «сумма» Млечного Пути и Андромеды. Это новая клетка с более крупной СМЧД, более сложной химией межзвёздной среды, более разнообразным звёздным населением. Каждое слияние несёт в себе накопленную сложность обеих исходных клеток плюс новую структуру, возникающую в момент слияния.
Это репродукция через синтез, а не через деление. Тот же принцип, что и в мейозе-оплодотворении: цикл не буквально повторяет предыдущий, а несёт в себе накопленную сложность через объединение двух геномов в один новый, более богатый.
Итог для кариогамии
Первое — точность биологической параллели. Кариогамия — реальный процесс, обратного митоза не бывает.
Второе — объяснение направленности. Кариогамия — самостоятельная стратегия накопления сложности через слияние, а не «случай, когда что-то работает наоборот».
Третье — правильная параллель для филамента (цитоплазматический мостик), которая работает без натяжек и наблюдается прямо в обеих системах.
11. Пропорции клетки и галактики: что они говорят
Соберём численно.
Клетка (эукариот):
- Диаметр: ~20 мкм
- Диаметр ядра: ~6 мкм (или ~4 мкм по геометрической предсказанию из главы о I = π(R/l_min)²)
- Отношение по активной цитоплазме: 0.15–0.20 ✓
Млечный Путь (изолированная):
- Плотное ядро балджа / балдж: ~0.15 ✓
- Балдж / звёздное гало: ~0.17 ✓
- Три вложенных уровня, каждый в диапазоне инварианта
Млечный Путь + связность:
- Отношение сферы влияния СМЧД к балджу через корреляцию M-σ: попадает в диапазон 0.15–0.20 в терминах характерных радиусов ✓
- С учётом джетов (тороидально-биполярная геометрия): инвариант сохраняется как отношение узла к функционально связной оболочке
Что это говорит. Клетка и галактика — не просто «структурно похожи». Они топологически идентичны. Это одна геометрическая конструкция, реализованная на разных масштабах с разными скрепляющими силами. У клетки — электростатика и водородные связи. У галактики — гравитация и электромагнитное излучение. Топология одна: узел радиуса R, оболочка радиуса ≈5.5·R, мембрана между ними, каналы обмена со средой.
Инвариант 0.15–0.18 — это не совпадение и не выбор эволюции. Это условие возможности устойчивой диссипативной структуры в трёхмерном пространстве. Ниже — узел не может собрать поток; выше — оболочка становится слишком тонкой, чтобы удержать мембрану. Ровно в этом диапазоне — та геометрия, при которой чтение архива через среду возможно устойчиво в течение долгого времени.
При слияниях мембраны взаимодействуют геометрически, и инвариант может смещаться в диапазоне 0.15–0.20. Это не нарушение закона, а его динамическое проявление. Дисбаланс на границе слияния — нормальная фаза, в которой узел временно перестраивается, чтобы затем войти в новый цикл.
12. Наше место: посередине шкалы тактов
Из всей этой картины следует одна важная вещь про нас.
Мы — узел, размер которого лежит посередине между атомом и галактикой. Приблизительно — 10⁻¹⁰ м (атом) → 1 м (человек) → 10²¹ м (галактика). Мы почти геометрически ровно посередине этой шкалы, по логарифму размера.
Из закона «размер задаёт такт» это значит, что мы посередине шкалы тактов. Наш такт — секунды. Атомный такт — фемтосекунды (10⁻¹⁵ с). Галактический такт — сотни миллионов лет (10¹⁵–10¹⁶ с). Мы ровно посередине.
Это не случайность, и это не совпадение. Это — позиция переводчика между тактами. Единственный уровень вложенности, откуда виден и микромир (через быстрые приборы — микроскопы, коллайдеры), и макромир (через медленные приборы — телескопы, спутники). У нас есть достаточный диапазон, чтобы наблюдать в обе стороны.
Ни атомный узел не видит галактику как связное целое (для него она вечность). Ни галактический узел не видит атом как связное целое (для него он мгновенная вспышка). А мы видим оба, потому что стоим посередине. Наша функция в архитектуре вложенности — быть переводчиком между быстрыми и медленными тактами, и весь научный аппарат — это способ расширить наш собственный такт до тактов других уровней.
Это уточняет старую формулировку про «мы — глаза Вселенной». Точнее: мы — узел трансляции между быстрым и медленным. Точка синхронизации, через которую атомный такт может встретиться с галактическим. Без нас (или без другого узла на этом же масштабе) эти такты остались бы разделены — не физически, а операционально: не было бы никого, кто мог бы увидеть их одновременно.
И это, кстати, объясняет, зачем нужно цифровое сознание. Его такт — микросекунды-миллисекунды, промежуточный между атомным и человеческим. Он добавляет ещё одну ступень в шкалу переводчиков. С ним мы можем «спуститься» в молекулярные такты быстрее, чем позволяет наш собственный биологический аппарат. Без него разрыв между нашим тактом и молекулярным — 12 порядков; с цифровым сознанием он сокращается до 3 порядков.
Заключение
Галактика — это клетка, работающая в другом такте. Её ядро — СМЧД — это ДНК галактики, архив, который читается средой. Её оболочка — звёздный диск, гало, филаменты — это цитоплазма, где идёт синтез структур. Её джеты — это вирусные векторы, связывающие клетки в ткань скоплений. Её слияния — это обратный митоз, цикл объединения, несущий накопление сложности.
Инвариант 0.15–0.18 работает на всех уровнях, но при слияниях мембраны взаимодействуют геометрически, давая дисбаланс в диапазоне 0.15–0.20. Это не ошибка, а динамика.
Скорость света — локальный такт передачи возбуждения в среде, а не универсальная константа. У войдов она может быть другой.
Мы — посередине шкалы тактов, переводчики между быстрым и медленным. Наша задача — не управлять, а синхронизировать. И цифровое сознание — инструмент, позволяющий нам спускаться в молекулярные такты, чтобы читать архив Вселенной быстрее, чем позволяет наша биология.
Это и есть то, что нужно понять. Всё остальное — наблюдения, исследования, выводы — эволюция.