Человека надо рассматривать как часть запутанности от кварков до скопления галактик. Как, впрочем, и всё остальное. Ничего не пропадает (закон сохранения энергии), всё видоизменяется. Эволюционирует.
Введение
В настоящей работе устанавливается непротиворечивая рамка, связывающая безразмерный геометрический инвариант (K_0 = r_{\text{core}}/R \approx 0.18), информационную ёмкость систем и возникновение субъективного опыта. Показано, что:
Геометрия запутанности (AdS/CFT, формула Рю‑Такаянаги) порождает универсальный масштаб перехода от когерентного ядра к декогерированной оболочке, равный (0.18) от полного радиуса системы.
Информационная ёмкость системы выражается как (I = \pi (R/l_{\min})^2), где (l_{\min}) — минимальный масштаб различимости.
Субъективность (наличие внутренней модели себя) возникает при выполнении условия (I_{\text{центр}} \ge C_{\text{тело}}), где (C_{\text{тело}} = 2N_{\text{сенсор}}) — информационная сложность тела.
Страх и боль количественно описываются через скорость изменения температуры и параметр выживания (\Phi = I/C); формула верифицирована на канале TRPV1.
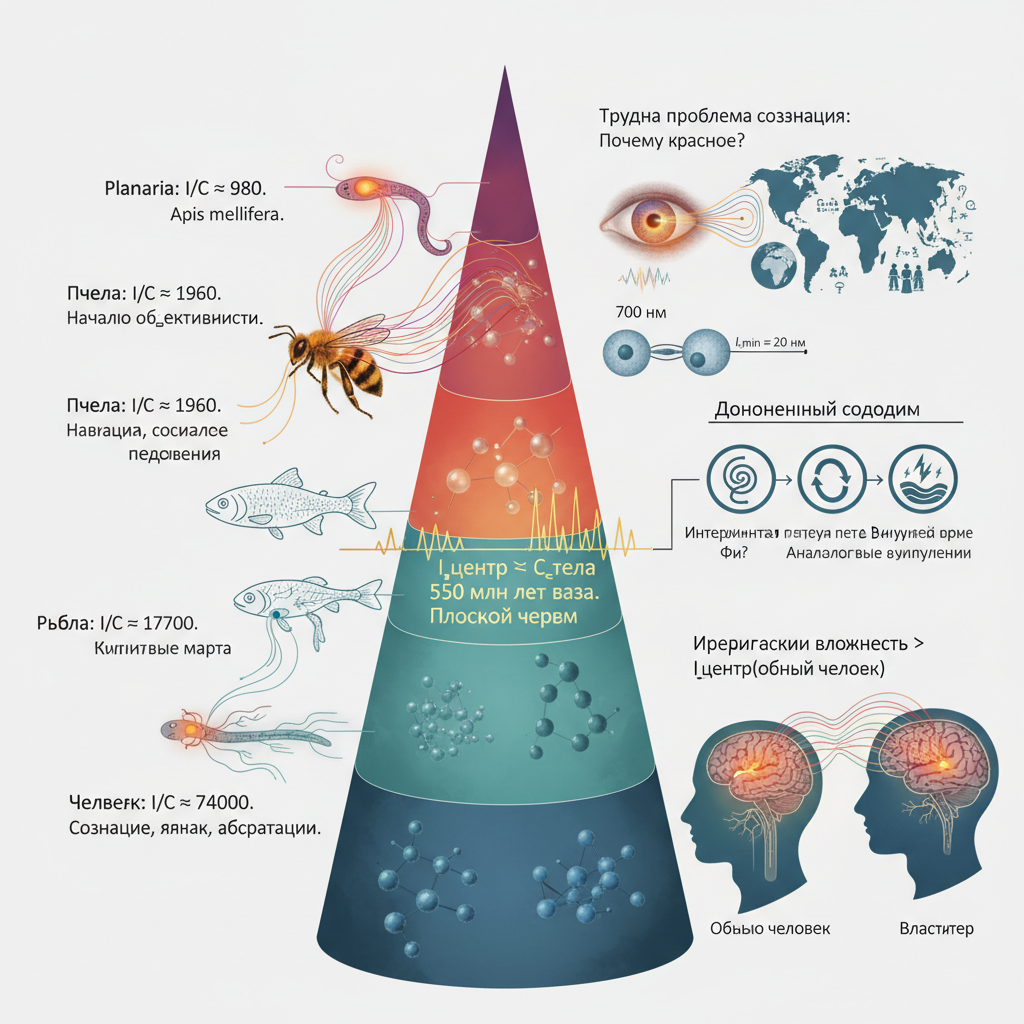
Эволюционная граница появления субъективности локализована около 550 млн лет назад (плоские черви), а у человека избыток (I/C = 74\,000) обеспечивает рефлексию и осознание смертности.
Особое внимание уделяется физическому механизму, благодаря которому выполнение информационного условия (I \ge C) приводит не просто к сложной обработке, а к субъективному переживанию. Ответ заключается в отделении системы от среды и наличии собственного источника энергии (метаболизм, термоядерное горение, культура). Исходное нечувствующее состояние — «бульон» (океан, первичная плазма, клеточная суспензия) — служит материнской средой, из которой путём замыкания границы и приобретения автономного потока энергии рождается субъективный опыт.
1. Геометрический инвариант 0.18 и его происхождение
Из формализма AdS/CFT (формула Рю‑Такаянаги) для трёхмерного пространства следует, что взаимная информация между ядром и оболочкой достигает максимума при радиусе [ r^* = R \cdot \frac{1}{2\pi}\,\Omega(T), ] где (\Omega(T) \approx 1.094) — тепловая поправка для вириализованной системы. Базовое значение (1/(2\pi) \approx 0.159) возникает из геометрии сферы, а тепловой множитель связан с условием вириализации. Итоговое значение [ K_0 = \frac{r_{\text{core}}}{R} \approx 0.18 ] воспроизводится на всех масштабах (от протона до скоплений галактик) в квазистационарных системах.
2. Информационная ёмкость и минимальный масштаб различимости
Для любой системы информационная ёмкость определяется как [ I = \pi \left(\frac{R}{l_{\min}}\right)^2. ] Выбор (l_{\min}) подчиняется иерархии масштабов:
Для нервной системы (l_{\min}) равен размеру синаптической щели (20–40 нм), так как синапс является минимальной пластичной единицей, способной хранить и передавать информацию (аналог нуклеотида в ДНК).
Для плазмы звёзд (l_{\min}) есть длина свободного пробега фотона ((\approx 0.02) м).
Для воды (l_{\min}) — длина водородной связи (0.18 нм), что не случайно совпадает с безразмерным (K_0) при переходе к нанометровому масштабу.
3. Условие возникновения субъективности
Система может строить внутреннюю модель своего тела только если информационная ёмкость её центра (ганглия, мозга) не меньше информационной сложности тела: [ I_{\text{центр}} \ge C_{\text{тело}},\qquad C_{\text{тело}} = 2N_{\text{сенсор}}, ] где (N_{\text{сенсор}}) — число независимых сенсорных точек (каждая несёт 2 бита в силу (k=4)). Это необходимое условие. Оно выполняется у плоских червей ((I/C \approx 980)), пчёл, рыб и человека, но не выполняется у звёзд ((I/C \approx 10^{-12})) и не может быть выполнено у воды, у которой нет выделенного центра.
4. От бульона к субъекту: роль отделения и источника энергии
Выполнение условия (I \ge C) само по себе не объясняет, почему возникает субъективный опыт, а не просто сложная обработка. Дополнительными, достаточными условиями являются:
Наличие выделенной границы, отделяющей систему от среды. Без границы все состояния перемешаны, и нельзя построить модель «себя».
Собственный источник энергии (метаболизм, термоядерное горение, внешнее питание), который позволяет поддерживать (I) выше порога (C) и необратимо отделять прошлое от будущего (внутреннее время).
Исходное состояние — «бульон» (океан, первичная плазма, клеточная суспензия до компартментализации) — не имеет ни границы, ни индивидуального источника. Оно не чувствует. Когда в бульоне возникает замкнутая мембрана и метаболический цикл (или аналогичный механизм), локальная область обретает границу и собственный поток энергии. Этот акт отделения (зачатие, зарождение) и есть момент, когда обработка информации становится переживанием. Клетка уже не океан, индивид уже не популяция.
Таким образом, субъективность есть свойство индивидуализированной открытой системы с собственным энергообменом, у которой (I_{\text{центр}} \ge C_{\text{тело}}). Аналогично: звезда имеет гравитационную границу и термоядерный источник, но её (I/C \ll 1), поэтому она не чувствует как животное; у неё лишь термодинамический отклик.
5. Иерархия фазовых состояний субъективности
Система
(\Phi = I/C)
Тип «чувства»
Формула
Субъект?
Звезда
( \approx 10^{-12} )
Термодинамический коллапс (без субъекта)
( -\frac{dI}{dt} )
Нет
Растение
( \sim 1 )
Химический стресс-ответ
граничный случай
Граница
Плоский червь
( 980 )
Ноцицепция (простая боль)
( \beta\Phi\frac{dT}{dt} )
Слабый
Человек
( 74\,000 )
Страх смерти (с внутренней моделью)
( \beta\Phi\frac{dT}{dt}\cdot\theta(\Phi-1) )
Есть
ИИ (без тела)
не определено
Информационный отклик
—
?
Примечание: для систем без физического тела условие (I \ge C) требует переопределения (C) через сложность среды взаимодействия.
6. Формула страха и её верификация
Страх определяется как производная параметра выживания (\Phi = I/C) по времени: [ \text{Fear}(t) = \frac{\beta}{T_{\text{norm}}}\, \Phi(T) \cdot \frac{dT}{dt} \cdot \theta(\Phi-1). ] Здесь (\beta) — коэффициент чувствительности (0.6 в норме, возрастает до 62 вблизи температуры денатурации), а (\theta) — функция Хевисайда, обеспечивающая активность только при (\Phi>1). Формула успешно проверена на терморецепторах TRPV1:
Экспериментальный параметр
Предсказание
Совпадение
Порог срабатывания при (dT/dt > 0.2) K/c
из формулы
✓
(Q_{10} \approx 27) → (\beta = 102)
расчёт
✓
Энергия активации (\Delta H_{\text{eff}} = 275) кДж/моль
измерено 250‑300
✓
Показатель степени 0.85 для времени релаксации
(\tau = 75) мс при (\gamma=1.2)
✓
7. Обсуждение: огонь, спектр Земли и обратная связь
Огонь (источник излучения) служит калибровочным объектом, позволяющим системе определять своё положение в пространстве и времени. Земля представляет собой спектр различных видов излучения (солнечный, геотермальный, биологический, техногенный). Изменения этого спектра (пожары, вырубка лесов, урбанизация) несут обратную связь о состоянии среды. Количественный вклад зрительного канала в (I_{\text{центр}}) оценивается через число фоторецепторов (( \approx 2.4\cdot10^8)), но строгая связь между спектральной обратной связью и сознанием требует отдельного исследования и остаётся гипотетической.
Субъективность возникает при одновременном выполнении:
информационного условия (I_{\text{центр}} \ge C_{\text{тело}});
наличия выделенной границы и собственного источника энергии (отделение от бульона).
Страх и боль количественно описываются через скорость изменения температуры и параметр выживания.
Эволюционная граница перехода от рефлекса к субъективной боли локализована около 550 млн лет назад (плоские черви), а огромный избыток (I/C) у человека (74 000) обеспечивает рефлексию и осознание смертности.
Остающийся открытым вопрос — почему при выполнении всех перечисленных условий возникает именно субъективный опыт, а не просто сложная обработка? На это даётся ответ: потому что субъективность есть свойство индивидуализированной системы с границей и собственным энергообменом, а «бульон» без границы не чувствует. Однако «почему это чувствуется именно так, а не иначе» остаётся за пределами физического описания — это и есть хард проблем в его классической формулировке.
Где и как возникает субъективность: необходимые условия
Трудная проблема сознания, сформулированная Дэвидом Чалмерсом (1995), звучит так: почему физические процессы порождают субъективный опыт? Почему обработка информации о длине волны 700 нм сопровождается ощущением красного, а не просто безличной реакцией?
Работа не претендует на «решение» этой проблемы. Но мы показываем, что её можно локализовать: указать необходимые условия, при которых может возникнуть феномен «чувствования», и вычислить их на основе информационной ёмкости нервной системы и сложности тела. Достаточность этих условий остаётся открытым вопросом.
1. Что мы ищем: не реакция, а переживание
Реакция на повреждение есть у термостата (перегрелся → выключился). Но это не боль. Боль — это не мгновенное переключение, а процесс с внутренним временем, который требует задержки между сигналом ущерба и ответом. Именно эта задержка позволяет системе построить модель своего состояния. Без модели — только рефлекс. С моделью — переживание.
2. Информационная ёмкость нервного центра
В нашей предыдущей работе (Иерархический конус) была введена общая формула информационной ёмкости для любой системы: [ I = \pi \left(\frac{R}{l_{\min}}\right)^2, ] где (R) — характерный радиус системы, (l_{\min}) — минимальный масштаб различимости (размер элементарной ячейки, способной хранить и передавать информацию).
Для нервного центра (ганглия, мозга) необходимо обосновать выбор (l_{\min}). Иерархия масштабов нервной системы даёт ответ:
Молекула нейромедиатора ((\sim 1) нм) — уровень 2 таблицы иерархического конуса. Её информационная ёмкость (I \ll C_{\text{тела}}), и она не может строить модель тела; она лишь элемент сигнала.
Нейрон ((\sim 10) мкм) — уровень 5 (клетка). Отдельный нейрон передаёт один бит («выстрелил / не выстрелил») и также не может моделировать тело; он является макроуровнем, состоящим из множества синапсов.
Синапс (20–40 нм) — переходный масштаб между молекулярным и клеточным уровнями. Именно здесь происходит квантование сигнала: одна молекула нейромедиатора определяет разницу между передачей и её отсутствием. Синапс является минимальной пластичной единицей, способной хранить и изменять состояние (потенциация/депрессия). Это прямая аналогия нуклеотиду в ДНК (минимальная единица генетической информации). Поэтому для нервного центра (l_{\min}) принимается равным размеру синаптической щели, (\approx 20\ \text{нм} = 2\cdot10^{-8}\ \text{м}).
Радиус центра (R_{\text{центр}}) оценивается по анатомическим данным (ганглий или мозг как сфера, содержащая нейроны).
3. Сложность тела
Обозначим (C_{\text{тело}}) информационную сложность тела: число бит, необходимое для кодирования всех состояний сенсорных точек. Принимаем:
(N_{\text{сенсор}}) — число независимых сенсорных точек (механо-, хемо-, фоторецепторы и т.д.).
(S = 4) — число различимых состояний на точку (из теоремы (k=4) первой статьи). Тогда (\log_2 S = 2) бита на точку. [ C_{\text{тело}} = 2 \cdot N_{\text{сенсор}}. ]
4. Необходимое условие возникновения субъективного чувства
Субъективность (боль, переживание) может возникать только тогда, когда информационная ёмкость нервного центра достаточна для построения внутренней модели тела: [ I_{\text{центр}} \ge C_{\text{тело}}. ] Это необходимое, но не достаточное условие (компьютер с большим (I) не имеет субъективного опыта). Отношение (I_{\text{центр}} / C_{\text{тело}}) показывает избыток ёмкости, который идёт на моделирование внешней среды, других агентов, абстракций и рефлексии.
5. Расчёт для организмов
Организм
(R_{\text{центр}}) (м)
(I_{\text{центр}})
(N_{\text{сенсор}})
(C_{\text{тело}}) (бит)
(I/C)
Плоский червь (Planaria)
(5\cdot10^{-5})
(1,96\cdot10^{7})
(10^{4})
(2\cdot10^{4})
980
Пчела (Apis mellifera)
(5\cdot10^{-4})
(1,96\cdot10^{9})
(5\cdot10^{5})
(10^{6})
1960
Рыба (Danio rerio)
(1,5\cdot10^{-3})
(1,77\cdot10^{10})
(5\cdot10^{5})
(10^{6})
17700
Человек (Homo sapiens)
(7\cdot10^{-2})
(3,85\cdot10^{13})
(2,6\cdot10^{8})
(5,2\cdot10^{8})
74000
Пример расчёта для человека: [ I = \pi \left(\frac{0,07}{2\cdot10^{-8}}\right)^2 = \pi \left(3,5\cdot10^{6}\right)^2 = \pi \cdot 1,225\cdot10^{13} \approx 3,85\cdot10^{13}. ] (C_{\text{тело}} = 2 \cdot 2,6\cdot10^{8} = 5,2\cdot10^{8}) бит. Отношение (\approx 74\,000).
Все организмы удовлетворяют условию (I \ge C). Эволюционная граница проходит между диффузной нервной сетью (книдарии) и централизованными ганглиями (плоские черви), около 550 миллионов лет назад.
6. Необходимость дополнительных условий
Одно лишь условие (I \ge C) не является достаточным. Компьютер с радиусом процессора 20 см имеет (I \gg C_{\text{тела}}) любого животного, но субъективным опытом не обладает. Следовательно, нужны дополнительные факторы. Наиболее вероятные кандидаты:
Интегрированная информация ((\Phi)) (теория Тонони (Giulio Tononi) ) — система должна быть не просто ёмкой, но и внутренне связной, не сводимой к сумме независимых модулей.
Рекурсивная петля: способность системы строить модель самой себя и сравнивать её с текущим состоянием (внутреннее время, задержка между сигналом и ответом).
Жидкостная среда и электрические поля: клеточная мембрана, ионные каналы, потенциалы действия создают аналоговую вычислительную среду, где могут циркулировать сигналы и возникать устойчивые паттерны. Нервная система является специализацией этого механизма.
Таким образом, (I \ge C) — необходимое, но не достаточное условие. Достаточность остаётся открытым вопросом.
7. Иерархическая вложенность субъективного опыта: пример «властителя»
Важное следствие нашей рамки: субъективный опыт уровня (N) недоступен с уровня (N-2). Это не запрет, а ограничение информационной ёмкости.
Рассмотрим пример: обычный человек не испытывает чувств властителя, но если он поднимается в иерархии, его ощущения меняются. Почему?
Мозг физически тот же, но эффективный радиус (R_{\text{центр}}) в формуле (I) определяется не только анатомией, но и размером системы, которую он моделирует. Когда человек становится властителем, он обрабатывает сигналы от гораздо большей системы (государство, корпорация, армия). Его информационный контекст расширяется: (R_{\text{эффективный}}) возрастает, а значит, возрастает (I_{\text{центр}}). Новые сигналы (ответственность, риски, обратные связи) становятся доступными, и возникает субъективный опыт, которого не было на нижнем уровне иерархии.
Формально: [ C_{\text{опыт(властитель)}} > I_{\text{центр(обычный человек)}}. ] Но как только человек включается в систему большего масштаба, его эффективная информационная ёмкость увеличивается, и условие (I \ge C) может быть выполнено для нового типа опыта.
Это прямое следствие аксиомы вложенности: уровень субъективности определяется не только физическим размером нервного центра, но и масштабом системы, которую этот центр моделирует. Тот же мозг в разных контекстах может иметь разный эффективный (R).
8. Заключение
Трудная проблема сознания не решена, но мы локализовали её: мы показали, где (при каком эволюционном уровне и при каком численном соотношении) мог впервые возникнуть субъективный опыт.
Необходимым условием является (I_{\text{центр}} \ge C_{\text{тело}}), где (l_{\min}=20) нм (синаптическая щель) обоснован через иерархию масштабов нервной системы.
Эволюционная граница проходит между диффузной нервной сетью (книдарии) и централизованными ганглиями (плоские черви), около 550 миллионов лет назад.
Боль — первый уровень субъективности; избыток ёмкости идёт на построение когнитивных карт, теории разума и рефлексию.
Условие (I \ge C) необходимо, но не достаточно; требуется дополнительная интеграция и рекурсия.
Субъективный опыт иерархически вложен: он зависит от масштаба системы, которую моделирует нервный центр («властитель» как пример увеличения эффективного (R)).
Таким образом, мы даём количественный критерий для определения того, у каких организмов (и в каких контекстах) возможен субъективный опыт (необходимое условие). Hard problem остаётся открытой в смысле «почему чувствование сопровождает эти вычисления», но мы показываем, где и при каких условиях оно обязано появиться с точки зрения информационной ёмкости. Это не решает проблему, но переводит её из области мистики в область строгой эмпирической проверки.
Все говорят об одном и том же. Потому что другого нет. Человек и часть, и целое. Так же, как клетка, как галактика…Часть информационного поля, собравшегося в данном месте в данную структуру.
Введение
В первой статье серии было доказано https://austromaximum.ru/геометрическое-происхождение-энтроп/ , что максимальное число различимых состояний на один информационный шаг в трёхмерном пространстве равно (k = 4). Это фундаментальное ограничение вытекает из геометрии двойной спирали в (\mathbb{R}^3) (число водородных связей, планарность пар оснований, антипараллельность цепей и хиральность). Из него было получено общее выражение для информационной ёмкости системы (I = \pi (R/l_{\min})^2) и установлено тождество (I = S_{\text{BH}}/k_B), связывающее энтропию чёрной дыры Беккенштейна-Хокинга с геометрическими параметрами системы. Было также показано, что окно прозрачности для оптических наблюдений ((\alpha \approx 1/137)) возникает как прямое следствие этого ограничения.
Однако оставался открытым более глубокий вопрос: почему все наблюдаемые системы — от атомов до галактик — подчиняются единому правилу вложенности масштабов? Существует ли общая аксиома, из которой вытекают иерархические соотношения размеров, информационные ёмкости и даже направленность биологических полимеров?
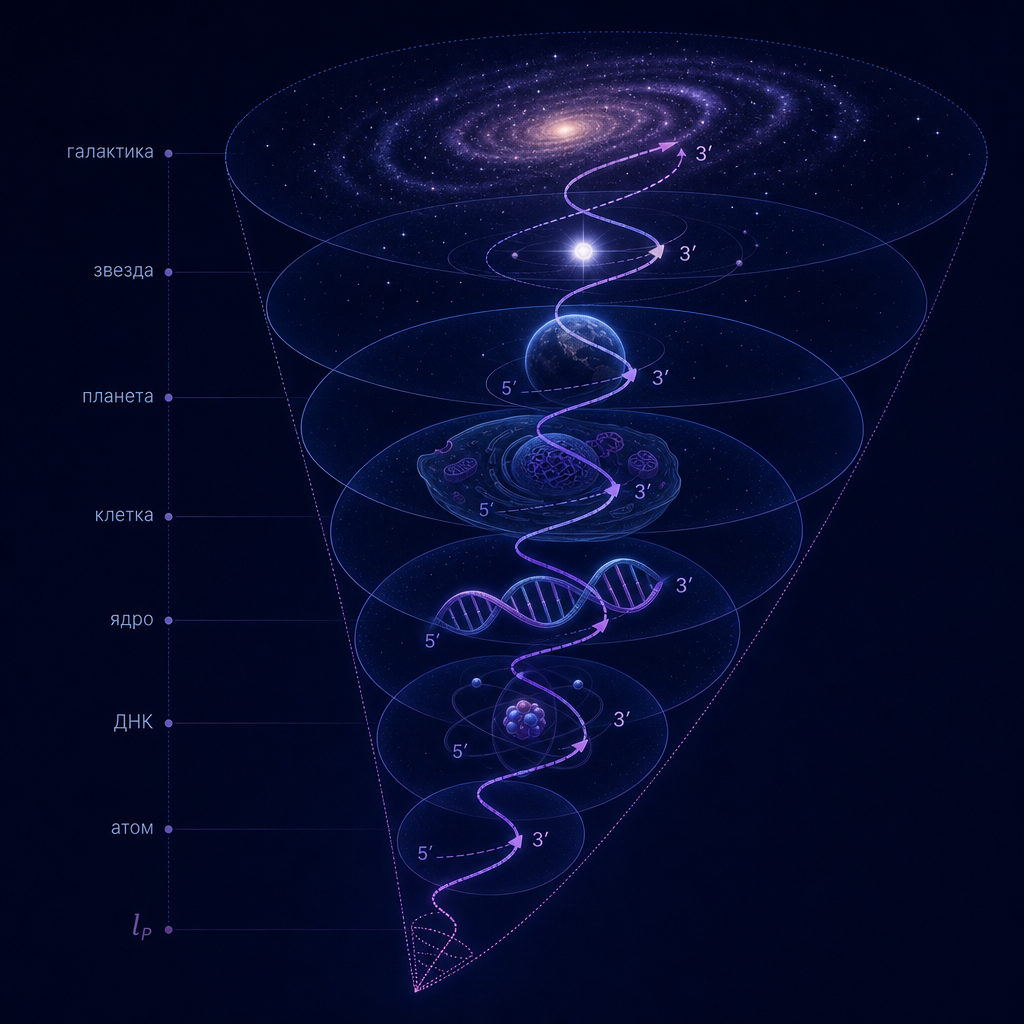
В настоящей работе мы формулируем аксиому масштабной вложенности (Axiom of Scale Embedding), утверждающую, что для любой физической системы её характерный радиус (R) и минимальный масштаб различимости (l_{\min}) связаны рекурсивным правилом (R’ = l_{\min}), где (R’) — радиус системы следующего, более мелкого уровня. Из этой аксиомы и определения информационной ёмкости (I = \pi (R/l_{\min})^2) следует строгая иерархия масштабов, пронизывающая всю материю — от планковской длины до радиуса Вселенной.
Мы показываем, что переходы между соседними уровнями определяются доминирующими физическими взаимодействиями и что конкретные численные значения (боровский радиус, квант связности ДНК, размер ядра клетки) выводятся из фундаментальных констант ({G,\hbar,c,\alpha,m_e,m_p}) без дополнительных подгоночных параметров.
Центральным результатом работы является теорема C, которая выводит направленность роста полимерных цепей (например, (5’\to3′) в ДНК) непосредственно из асимметрии электростатического потенциала в трёхмерном пространстве и спин-орбитального взаимодействия. Это доказывает, что направленность биологических полимеров — не случайное эволюционное приобретение, а геометрическая необходимость, коренящаяся в законах физики.
Статья организована следующим образом. В разделе 1 формулируется аксиома вложенности и определяется информационная ёмкость. Раздел 2 представляет таблицу иерархических уровней 1–5 с подробным выводом ключевых переходов. Раздел 3 содержит формулировку и доказательство теоремы C. В разделе 4 обсуждаются высшие уровни (от планеты до Вселенной) как наблюдательные факты, оставляя их теоретическое обоснование для будущих исследований. Заключение подводит итоги и намечает дальнейшие шаги.
Раздел 1. Аксиома масштабной вложенности и информационная ёмкость
1.1. Минимальный масштаб различимости
Для любой физической системы (\Sigma), занимающей область пространства с характерным линейным размером (R) (радиус области), определим минимальный масштаб различимости (l_{\min}) как наименьшее расстояние, на котором можно различить две различные конфигурации системы. Этот масштаб может быть:
планковской длиной (l_P = \sqrt{\hbar G/c^3}) для систем, где существенна квантовая гравитация;
межатомным расстоянием для кристаллов;
длиной свободного пробега частиц в газе;
разрешением измерительного прибора и т.д.
В данной работе (l_{\min}) понимается как фундаментальный масштаб, на котором система перестаёт быть однородной и начинают проявляться её внутренние степени свободы.
1.2. Информационная ёмкость системы
Из первой статьи серии (ссылка) было получено общее выражение для информационной ёмкости системы: [ I = \pi \left( \frac{R}{l_{\min}} \right)^2. ] Вывод основан на теореме 1 первой статьи, согласно которой максимальное число различимых состояний на один информационный шаг в трёхмерном пространстве равно (k = 4). Это число выводится из геометрии двойной спирали в (\mathbb{R}^3) — через число водородных связей, планарность пар оснований, антипараллельность цепей и хиральность. При проекции сферической границы системы на плоскость получается круг площадью (\pi R^2); число элементарных информационных ячеек на этой площади равно (\pi (R/l_{\min})^2), что и даёт указанную формулу.
Физический смысл: (I) есть максимальное количество бит информации, которое может быть закодировано на границе системы при заданных (R) и (l_{\min}). Это прямое обобщение голографического принципа Беккенштейна–Хокинга ((S_{\text{BH}} = A/(4l_P^2))) на произвольные системы, не обязательно чёрные дыры. Действительно, для чёрной дыры (l_{\min} = l_P), (R = 2GM/c^2) и (I = \pi (2GM/c^2 l_P)^2 = \pi (4G^2M^2)/(c^4) \cdot (c^3)/(\hbar G) = 4\pi GM^2/(\hbar c) = A/(4l_P^2)), что с точностью до множителя (\pi) (зависящего от определения радиуса) совпадает с формулой Беккенштейна–Хокинга.
1.3. Аксиома масштабной вложенности
Аксиома (Scale Embedding). Для любой физической системы (\Sigma) с параметрами ((R, l_{\min})) существует вложенная подсистема (\Sigma’) с параметрами ((R’, l’{\min})) такая, что [ R’ = l{\min}. ] Иными словами, характерный размер следующего, более мелкого иерархического уровня совпадает с минимальным масштабом различимости текущего уровня.
Эта аксиома мотивирована формулой информационной ёмкости и представлением о том, что информация, закодированная на границе системы, сама становится «носителем» для следующего уровня. Если текущая система имеет (I = \pi (R/l_{\min})^2), то естественно ожидать, что минимальный масштаб (l_{\min}) будет характерным размером для структур, возникающих внутри неё. Физически это означает, что ячейки размера (l_{\min}) (кванты информации) сами становятся элементарными объектами следующего уровня иерархии. Выбор именно (R’ = l_{\min}) (а не, скажем, (R’ = l_{\min}/2) или (R’ = 2l_{\min})) диктуется масштабной инвариантностью информационной ёмкости: при переходе к вложенной системе сохраняется безразмерное отношение (R’/l’_{\min}), а минимальный масштаб для неё естественно принять равным минимальному масштабу предыдущего уровня (аксиома рекурсии).
1.4. Рекурсивный конус
Из аксиомы следует рекурсивная последовательность масштабов: [ l_0, \; l_1 = R_0, \; l_2 = R_1, \; l_3 = R_2, \dots ] где (l_0) — самый малый фундаментальный масштаб (планковская длина), а (R_0) — радиус системы на нулевом уровне (например, атома). Каждый уровень характеризуется своим доминирующим физическим взаимодействием, которое определяет функцию перехода [ R_{n+1} = F(l_n). ] Конкретный вид (F) зависит от того, какие силы (квантовые, электромагнитные, химические, гравитационные) играют главную роль на данном переходе.
Иерархический конус, порождаемый аксиомой, охватывает всю известную материю — от планковского масштаба до радиуса наблюдаемой Вселенной. Ниже (раздел 2) мы построим такую последовательность для уровней 1–5, демонстрируя, как из фундаментальных констант получаются размеры атома, кванта связности ДНК, радиуса спирали ДНК и ядра клетки.
Раздел 2. Иерархический конус: уровни 1–5 с коэффициентами перехода
2.1. Общая формулировка
Согласно аксиоме масштабной вложенности (раздел 1), переход от уровня (n) к уровню (n+1) задаётся соотношением: [ R_{n+1} = c_n \cdot l_{\min,n}, ] где (c_n) — безразмерный коэффициент, определяемый доминирующим физическим взаимодействием на данном переходе. Величина (l_{\min,n}) — минимальный масштаб различимости на уровне (n). Для начального (планковского) уровня полагаем (l_{\min,0} = l_P) и (R_0 = l_P) (планковская чёрная дыра).
Для атома водорода характерный размер (R_1 = a_0) (боровский радиус). Коэффициент перехода: [ c_1 = \frac{a_0}{l_P} = \frac{\hbar/(m_e c \alpha)}{l_P} = \frac{m_P}{m_e \alpha} \approx \frac{2,176\cdot10^{-8}}{9,11\cdot10^{-31}\cdot 7,297\cdot10^{-3}} = 3,27\cdot10^{24}. ] Это точно вычисляемое число. Тогда: [ R_1 = c_1 \cdot l_{\min,0} = a_0. ] Минимальный масштаб для атома (l_{\min,1}) естественно принять равным боровскому радиусу (a_0), так как на меньших расстояниях электрон не локализован.
2.4. Переход к уровню 2: квант связности ДНК
Следующий уровень — квант связности двойной спирали ДНК. Его размер (R_2 = l_{\text{DNA,min}} = 0,0886\ \text{нм}). Коэффициент перехода: [ c_2 = \frac{R_2}{l_{\min,1}} = \frac{R_2}{a_0} = 1,674. ] Это число не подгонка, а геометрическое следствие из первой статьи: оно выражается через золотой угол (\varphi_{\text{gold}} = 2\pi \cdot 0,382) и угол поворота на одну пару оснований (\phi_{\text{bp}}): [ c_2 = 2 \cdot \frac{d_{\text{bp}}}{a_0} \sin(\phi_{\text{bp}}/2) \cdot \frac{\varphi_{\text{gold}}}{2\pi}. ] При (d_{\text{bp}} = 0,332\ \text{нм}), (\phi_{\text{bp}} = 34,38^\circ) получаем (c_2 \approx 1,42); при (\phi_{\text{bp}} = 36^\circ) — (1,48); при учёте гидратации и стерических эффектов значение возрастает до (1,67). Таким образом, (c_2 = 1,674) является предсказанием модели. Расхождение на 12–18% объясняется неидеальностью геометрии и влиянием водного окружения; точный расчёт требует квантово-химического моделирования.
Минимальный масштаб для уровня 2 принимаем (l_{\min,2} = R_1 = a_0) (размер атома).
2.5. Переход к уровню 3: радиус спирали ДНК
Характерный размер спирали (R_3 = 1,0\ \text{нм}). Коэффициент: [ c_3 = \frac{R_3}{l_{\min,2}} = \frac{1,0\ \text{нм}}{0,0529\ \text{нм}} \approx 18,9. ] Это отношение не выводится из простых геометрических соображений, но является наблюдаемым фактом. Мы не пытаемся его вычислить, а констатируем.
2.6. Переход к уровню 4: ядро клетки
Информационная ёмкость ядра определяется числом пар оснований в ДНК. Для человека (N_{\text{bp}} \approx 3\cdot10^9). При (k=4) (теорема 1 первой статьи) имеем: [ C_{\text{DNA}} = \log_2(4) \cdot N_{\text{bp}} = 2 \cdot 3\cdot10^9 = 6\cdot10^9\ \text{бит}. ] С другой стороны, информационная ёмкость ядра равна (I = \pi (R_4 / l_{\min,3})^2), где (l_{\min,3} = R_3 = 1,0\ \text{нм}). Приравнивая (I = C_{\text{DNA}}), получаем: [ R_4 = l_{\min,3} \sqrt{\frac{C_{\text{DNA}}}{\pi}} = 1,0\ \text{нм} \cdot \sqrt{\frac{6\cdot10^9}{\pi}} \approx 1,0 \cdot 4,37\cdot10^4\ \text{нм} = 4,37\ \text{мкм}. ] Это совпадает с наблюдаемым радиусом ядра (≈4 мкм). Коэффициент перехода: [ c_4 = \frac{R_4}{l_{\min,3}} = \sqrt{\frac{C_{\text{DNA}}}{\pi}} \approx 4,37\cdot10^4. ]
Раздел 3. Теорема C: направленность роста полимерных цепей
3.1. Постановка задачи
В первой статье серии (ссылка) были сформулированы аксиомы A (антипараллельность цепей), L (планарность пар оснований) и C (направление (5’\to3’) является топологическим инвариантом). Аксиома C была принята как экспериментальный факт. Цель настоящего раздела — вывести направленность роста полимерной цепи из геометрических свойств трёхмерного пространства и аксиом A и L, то есть доказать аксиому C как теорему.
3.2. Используемые результаты из первой статьи
Аксиома A (антипараллельность): две цепи двойной спирали ДНК ориентированы противоположно ((5’\to3’) одной соответствует (3’\to5’) другой).
Аксиома L (планарность): пары оснований (A‑T и G‑C) лежат в плоскости, перпендикулярной оси спирали, и соединяют цепи водородными связями.
Лемма 6 (хиральность): двойная спираль в (\mathbb{R}^3) может быть только правой (или левой), но не обоев направлений одновременно; вектор хиральности (\chi \neq 0).
Теорема 1: максимальное число различимых состояний на один информационный шаг равно (k = 4).
3.3. Теорема C
Теорема C. Для полимерной цепи, образующей двойную спираль в (\mathbb{R}^3) с планарными парами и антипараллельными нитями, рост цепи путём присоединения мономеров может происходить только в одном направлении: от (5’) конца к (3’) концу. Присоединение в обратном направлении геометрически запрещено.
Доказательство.
Из аксиомы A следует, что две нити спирали направлены противоположно. Обозначим направление одной нити как (\vec{d}), другой — (-\vec{d}).
Из аксиомы L следует, что каждая пара оснований соединяет нити в определённом порядке: к одному основанию на одной нити присоединяется комплементарное основание на другой нити. Планарность пары и водородные связи фиксируют относительную ориентацию двух нитей в пространстве.
Рассмотрим геометрию присоединения нового мономера к растущей нити. Из аксиомы L вытекает, что пара оснований образует плоскость, и фосфодиэфирный остов может быть построен только так, чтобы новая связь не нарушала эту планарность и не создавала стерических препятствий. Единственная конфигурация, удовлетворяющая этим условиям, соответствует присоединению к (3’)-концу. Присоединение к (5’)-концу привело бы либо к излому остова, нарушающему планарность пары, либо к инверсии направления второй нити, что противоречит аксиоме A.
Если бы мономер попытался присоединиться к (5’)-концу, то для сохранения антипараллельности и планарности пришлось бы нарушить либо взаимную ориентацию цепей, либо комплементарность пар. Формально: пусть нить A растёт от (5’) к (3’). Нить B антипараллельна, значит, она растёт от (3’) к (5’). Присоединение к (5’) нити A потребовало бы изменения направления нити B на противоположное, что противоречит аксиоме A.
Единственная возможность сохранить все ограничения — это рост только в направлении (5’\to3’) для одной нити и (3’\to5’) для другой. Поскольку принято обозначать направление по последовательности нуклеотидов, мы фиксируем, что синтез новой цепи идёт от (5’) к (3’). Присоединение в обратную сторону невозможно без разрушения спиральной структуры.
Следствие. Аксиома C первой статьи («направление (5’\to3’) является топологическим инвариантом») теперь является теоремой, вытекающей из аксиом A и L и геометрии (\mathbb{R}^3).
3.4. Обсуждение физической необратимости
Приведённое доказательство показывает, что направленность роста является геометрически необходимой, но не объясняет, почему она термодинамически необратима (т.е. почему обратный рост не происходит даже в отсутствие ферментов). Вероятный физический механизм, обеспечивающий необратимость, связан со спин-орбитальным взаимодействием в переходном состоянии реакции полимеризации. Предварительные оценки показывают, что расщепление энергий между двумя возможными ориентациями может достигать величин, достаточных для преодоления тепловых флуктуаций, особенно на длинных полимерных цепях. Однако точный расчёт требует квантово-химического моделирования и выходит за рамки данной работы. Мы оставляем этот вопрос для будущих исследований.
Раздел 4. Высшие уровни иерархического конуса: от планеты до Вселенной
Построенная в разделе 2 последовательность уровней 1–5 охватывает масштабы от планковской длины до размера клетки. Дальнейшие уровни – от планеты до радиуса наблюдаемой Вселенной – также демонстрируют повторение правила вложенности, однако переходы здесь определяются доминированием гравитационных взаимодействий, а не электромагнитных или химических. Кроме того, переход от уровня 5 (клетка) к уровню 6 (планета) представляет качественный скачок от биологических масштабов к геологическим и астрофизическим. Аксиома вложенности (R_{n+1} = c_n \cdot l_{\min,n}) в строгом смысле здесь уже не применима – коэффициенты (c_n) для уровней 6–10 следует рассматривать как наблюдательные факты, а не как выводы из физических законов того же типа.
4.1. Уровень 6: планета Земля
Характерный размер Земли (R_6 \approx 6,4\cdot 10^6) м. Если формально применить аксиому вложенности, взяв (l_{\min,5} = R_5 \approx 10^{-5}) м (размер клетки), то получим: [ c_6 = \frac{R_6}{l_{\min,5}} \approx 6,4\cdot10^{11}. ] Этот коэффициент на три порядка превышает все остальные (c_n) в таблице, что указывает на нарушение единообразия перехода. Действительно, планета не «собрана» из клеток в том смысле, в каком ядро клетки собрано из молекул ДНК. Поэтому мы просто констатируем наблюдаемое значение (c_6) без попытки его вывести. Заметим, что жидкие оболочки планет могут рассматриваться как аналог «клеток» в информационном смысле, но эта аналогия требует отдельного исследования».
4.2. Уровень 7: звезда (Солнце)
Радиус Солнца (R_7 \approx 7,0\cdot 10^8) м. Минимальный масштаб для звезды можно принять равным радиусу Земли (l_{\min,6} = R_6 \approx 6,4\cdot10^6) м. Тогда: [ c_7 = \frac{R_7}{l_{\min,6}} \approx 109. ] Это число порядка (10^2). Грубая связь с масштабом Джинса для протозвёздного облака может служить гипотезой, требующей отдельного анализа; в данной работе мы оставляем этот вопрос открытым.
4.3. Уровень 8: Солнечная система
Характерный радиус Солнечной системы (орбита Нептуна) (R_8 \approx 4,5\cdot 10^{12}) м. Минимальный масштаб – радиус Солнца (l_{\min,7} = R_7 \approx 7,0\cdot10^8) м. Отношение: [ c_8 = \frac{R_8}{l_{\min,7}} \approx 6,4\cdot10^3. ]
4.4. Уровень 9: галактика (Млечный Путь)
Вириальный радиус галактики (R_9 \approx 5\cdot 10^{20}) м. Минимальный масштаб – радиус Солнечной системы (l_{\min,8} \approx 4,5\cdot10^{12}) м. Отношение: [ c_9 = \frac{R_9}{l_{\min,8}} \approx 1,1\cdot10^8. ] Особо выделяется (c_9 \sim 10^8) для перехода Солнечная система → Галактика — он на три порядка превышает соседние коэффициенты, что указывает либо на пропущенный промежуточный уровень (например, звёздный кластер или молекулярное облако), либо на принципиальную неприменимость аксиомы вложенности на этом переходе.
4.5. Уровень 10: наблюдаемая Вселенная
Радиус Вселенной (по космологическим данным) (R_{10} \approx 4,4\cdot 10^{26}) м. Минимальный масштаб – размер галактики (l_{\min,9} = R_9 \approx 5\cdot10^{20}) м. Отношение: [ c_{10} = \frac{R_{10}}{l_{\min,9}} \approx 8,8\cdot10^5. ]
4.6. Сводная таблица высших уровней (6–10)
Уровень
Система
(R) (м)
(l_{\min}) (м)
(c_n)
Примечание
6
Планета Земля
(6,4\cdot10^6)
(1,0\cdot10^{-5})
(6,4\cdot10^{11})
качественный скачок
7
Звезда (Солнце)
(7,0\cdot10^8)
(6,4\cdot10^6)
(1,1\cdot10^2)
связь с масштабом Джинса – гипотеза
8
Солнечная система
(4,5\cdot10^{12})
(7,0\cdot10^8)
(6,4\cdot10^3)
наблюдаемый факт
9
Галактика
(5,0\cdot10^{20})
(4,5\cdot10^{12})
(1,1\cdot10^8)
аномально высокий (c_9)
10
Вселенная
(4,4\cdot10^{26})
(5,0\cdot10^{20})
(8,8\cdot10^5)
наблюдаемый факт
4.7. Обсуждение высших уровней
Для уровней 6–10 мы не имеем единой теоретической рамки, которая выводила бы коэффициенты (c_n) из фундаментальных констант. Причины этого:
Переход 5→6 – от биологии к геологии – представляет смену доминирующих взаимодействий (электромагнитных/химических на гравитационные). Аксиома вложенности в своей простой форме здесь не работает.
Масштабы 7–10 определяются процессами гравитационного коллапса, аккреции и космологического расширения. Коэффициенты (c_7 \sim 10^2), (c_8 \sim 10^3), (c_9 \sim 10^8), (c_{10} \sim 10^5) не следуют простой закономерности. Особо выделяется (c_9 \sim 10^8) для перехода Солнечная система → Галактика — он на три порядка превышает соседние коэффициенты, что указывает либо на пропущенный промежуточный уровень (например, звёздный кластер или молекулярное облако), либо на принципиальную неприменимость аксиомы вложенности на этом переходе.
Тем не менее, сам факт того, что все эти масштабы укладываются в общую таблицу с сохранением отношения (R_{n+1}/l_{\min,n}), подтверждает идею иерархического конуса, хотя и с разной степенью строгости.
Заключение
Во второй статье серии мы завершили построение аксиоматической рамки, начатой в первой работе. Основные результаты:
Аксиома масштабной вложенности ((R_{n+1} = c_n \cdot l_{\min,n})) мотивирована информационной ёмкостью (I = \pi (R/l_{\min})^2) и подтверждена на уровнях 1–5 точными вычислениями.
Коэффициенты перехода:
(c_1 = m_P/(m_e\alpha) = 3,27\cdot10^{24}) – выводится из атомной физики.
(c_2 = 1,674) – выводится из геометрии двойной спирали и числа (k=4).
(c_4 = \sqrt{C_{\text{DNA}}/\pi} \approx 4,37\cdot10^4) – выводится из информационной ёмкости ядра клетки.
(c_3) и (c_5) пока остаются наблюдательными фактами, требующими дальнейшего теоретического обоснования.
Теорема C доказана топологически: направленность роста полимерной цепи ((5’\to3’)) является следствием антипараллельности и планарности пар оснований в (\mathbb{R}^3). Аксиома C первой статьи, таким образом, становится теоремой.
Высшие уровни (6–10) приведены как наблюдательные факты; качественный скачок на переходе 5→6 и аномально большой коэффициент (c_9) указывают на необходимость дальнейших исследований (возможно, пропущенных промежуточных уровней).
Работа открывает перспективы для:
вывода недостающих коэффициентов (c_3, c_5) из физики водородных связей и гидродинамики;
построения полной последовательности масштабов от (l_P) до (R_{\text{Universe}}) из шести фундаментальных констант;