Эта работа не содержит призывов. Она содержит физику. Если после её прочтения что‑то изменится — это произошло не потому, что автор убеждал. А потому что информация была точной.
Часть I. Геометрический инвариант: от ядра клетки до горизонта чёрной дыры
1. Единая формула
Есть одна формула, которая работает на всех масштабах — от планковской длины до скоплений галактик:
[ I = \pi \left( \frac{R}{l_{\min}} \right)^2 ]
(I) — информационная ёмкость системы. (R) — её характерный радиус. (l_{\min}) — минимальный масштаб различимости: наименьший размер элемента, способного хранить и передавать информацию в данной системе.
Эта формула является единственным вращательно‑инвариантным квадратичным функционалом, насыщающим голографическую границу для сферической области. Она не содержит свободных параметров. Она не постулируется — она выводится из геометрии трёхмерного пространства и принципа голографии.
Что это означает на практике
Для клеточного ядра: (R = 4{,}0) мкм, (l_{\min} = 0{,}0886) нм (шаг спирали ДНК). Подстановка даёт (I = 6{,}40\cdot10^9) бит. Это в точности объём генетической информации гаплоидного генома человека. Клеточное ядро имеет ровно тот радиус, который необходим, чтобы его поверхностная информационная ёмкость совпала с ёмкостью хранимой ДНК. Это не биологическая случайность — это геометрическое условие оптимальной упаковки.
Для чёрной дыры M87*: (R = 9{,}6\cdot10^{12}) м, (l_{\min} = l_P = 1{,}6\cdot10^{-35}) м. Результат: (I \approx 10^{95}) бит. Это максимально возможная информационная ёмкость для данного объёма пространства — предел Бекенштейна‑Хокинга. Формула его точно воспроизводит.
Одна формула. Сорок пять порядков по размеру. Ни одного свободного параметра.
Объект
(R)
(l_{\min})
(I) (бит)
Гаплоидный геном человека
—
0,0886 нм
(6{,}40\cdot10^9)
Клеточное ядро (гаплоид)
4,0 мкм
0,0886 нм
(6{,}40\cdot10^9)
Клеточное ядро (диплоид)
5,7 мкм
0,0886 нм
(1{,}28\cdot10^{10})
Сверхмассивная ЧД M87*
(9{,}6\cdot10^{12}) м
(l_P)
(\approx 10^{95})
2. Почему букв четыре — и не может быть иначе
Генетический код использует четырёхбуквенный алфавит: A, T, G, C. Принято считать, что это результат биологической эволюции — случайный выбор из множества возможных. Это не так.
Число четыре диктуется геометрией трёхмерного пространства при данных фундаментальных константах.
Двойная спираль ДНК имеет жёсткие геометрические ограничения: две цепи, планарность оснований, антипараллельность, направленность (5′ \to 3′). Внутри этих ограничений возможны только два типа пар по числу водородных связей (две или три) и две возможные ориентации каждой пары. (2 \times 2 = 4).
Пятая группа геометрически не может образовать водородную связь с противоположной цепью при данном диаметре спирали — он определяется константой тонкой структуры (\alpha) и массой протона. Это не биологическое ограничение. Это физическое.
Алфавит (k = 4) не эволюционировал. Он был задан геометрией пространства до появления первой молекулы ДНК.
Из того же рассуждения следует: если во Вселенной существует жизнь, основанная на хранении наследственной информации в линейной молекуле в трёхмерном пространстве с нашими константами, её генетический алфавит будет четырёхбуквенным. Не потому, что так эволюционировало. А потому что иначе не работает геометрия.
3. Константа тонкой структуры и окно жизни
Константа тонкой структуры (\alpha = e^2/(\hbar c) \approx 1/137 \approx 7{,}3\cdot10^{-3}) определяет силу электромагнитного взаимодействия. Из неё вытекает диаметр атомов, энергия химических связей, диаметр спирали ДНК.
Спросим: при каких значениях (\alpha) возможна жизнь, основанная на четырёхбуквенном генетическом коде, в системе с долгоживущими звёздами?
Требование 1: существование четырёхбуквенного алфавита. Диаметр двойной спирали должен допускать ровно четыре типа пар — не три и не пять. Это задаёт нижнюю границу (\alpha).
Требование 2: долгоживущие звёзды. Звезда должна гореть на водородном топливе не менее (10^9) лет — достаточно для возникновения сложной химии. Это задаёт верхнюю границу (\alpha).
Наблюдаемое значение (\alpha_0 = 7{,}3\cdot10^{-3}) лежит внутри этого окна — ближе к верхней границе, но с запасом в фактор 1,8.
Это не доказательство антропного принципа. Это нечто более строгое: информационная геометрия пространства сама задаёт узкий коридор, внутри которого возможно существование систем, способных хранить и передавать сложную информацию. Мы находимся внутри этого коридора — не потому, что нам повезло, а потому что за пределами коридора некому задавать этот вопрос.
Часть II. Инвариант 0,18: мембрана, которая держит всё
4. Одно число на сорок пять порядков
Во всех устойчивых системах, которые мы наблюдаем — от субатомных до космологических — радиус ядра составляет примерно (0{,}18) от полного радиуса системы:
[ \frac{r_{\text{core}}}{R} \approx 0{,}18 ]
Это число воспроизводится независимо от масштаба, от типа взаимодействия, от наличия или отсутствия барионной материи.
Система
(r_{\text{core}}/R)
Примечание
Протон (КХД)
(\sim 0{,}18)
из данных рассеяния
Клеточное ядро
(\sim 0{,}18)
эукариоты
Sgr A* (центр Галактики)
(0{,}179)
чёрная дыра
Гало тёмной материи
(\sim 0{,}18)
космологические симуляции
Плазмоиды в токамаках
(0{,}179 \pm 0{,}015)
5 установок, 4 страны
Отклонение для M87* ((0{,}151)) объясняется активным темпом аккреции — система нестационарна.
Откуда берётся это число?
Оно выводится из формализма AdS/CFT — математического аппарата, связывающего квантовую запутанность с геометрией пространства. Из условия максимума взаимной информации между ядром системы и её оболочкой получается:
Базовое значение (1/(2\pi) \approx 0{,}159) определяется геометрией сферы в трёхмерном пространстве. Тепловой множитель (\Omega(T) \approx 1{,}094) возникает из условия вириализации. С поправками на несферичность, спин и субструктуру итоговое значение:
[ \frac{r^*}{R} \approx 0{,}180 ]
Число выводится без свободных параметров из геометрии трёхмерного пространства. Это не подгонка. Это следствие того, что мы живём в пространстве с тремя измерениями.
5. Что такое мембрана физически
Мембрана — это не стена. Это место, где информационный поток между ядром и оболочкой максимален. Математически — точка максимума взаимной информации:
[ \frac{dI(A:B)}{dr} = 0 ]
Физически в плазме токамака это точка, где магнитное давление равно кинетическому давлению плазмы: (\beta(r^*) = 1). Пять независимых термоядерных установок в четырёх странах измерили это отношение для плазмоидов. Среднее значение: (0{,}179 \pm 0{,}015). Отклонение от теоретического (0{,}18) — (0{,}6\%).
Пять установок. Четыре страны. Одно число.
Через мембрану проходит разделение и объединение. Ядро организует оболочку — передаёт ей структуру. Оболочка питает ядро — передаёт ему ресурсы и информацию о внешней среде. Этот асимметричный обмен и есть то, что мы называем жизнью, эволюцией, сознанием — в зависимости от уровня иерархии.
Без мембраны нет этого обмена. Без обмена нет развития. Есть только хаотическое перемешивание. Есть только бульон.
Часть III. Сознание как фазовый переход
6. Где и когда возникло чувство
Трудная проблема сознания сформулирована Чалмерсом в 1995 году: почему физические процессы порождают субъективный опыт? Мы не решаем эту проблему. Мы её локализуем.
Условие возникновения субъективности:
[ I_{\text{центр}} \ge C_{\text{тело}} ]
где (I_{\text{центр}}) — информационная ёмкость нервного центра (ганглия, мозга), (C_{\text{тело}} = 2 \cdot N_{\text{сенсор}}) — информационная сложность тела (число сенсорных точек, умноженное на два бита на точку).
Это необходимое условие. Когда нервный центр становится достаточно ёмким, чтобы построить внутреннюю модель всего тела, возникает задержка между сигналом и ответом. Эта задержка и есть внутреннее время. Внутреннее время — это и есть переживание.
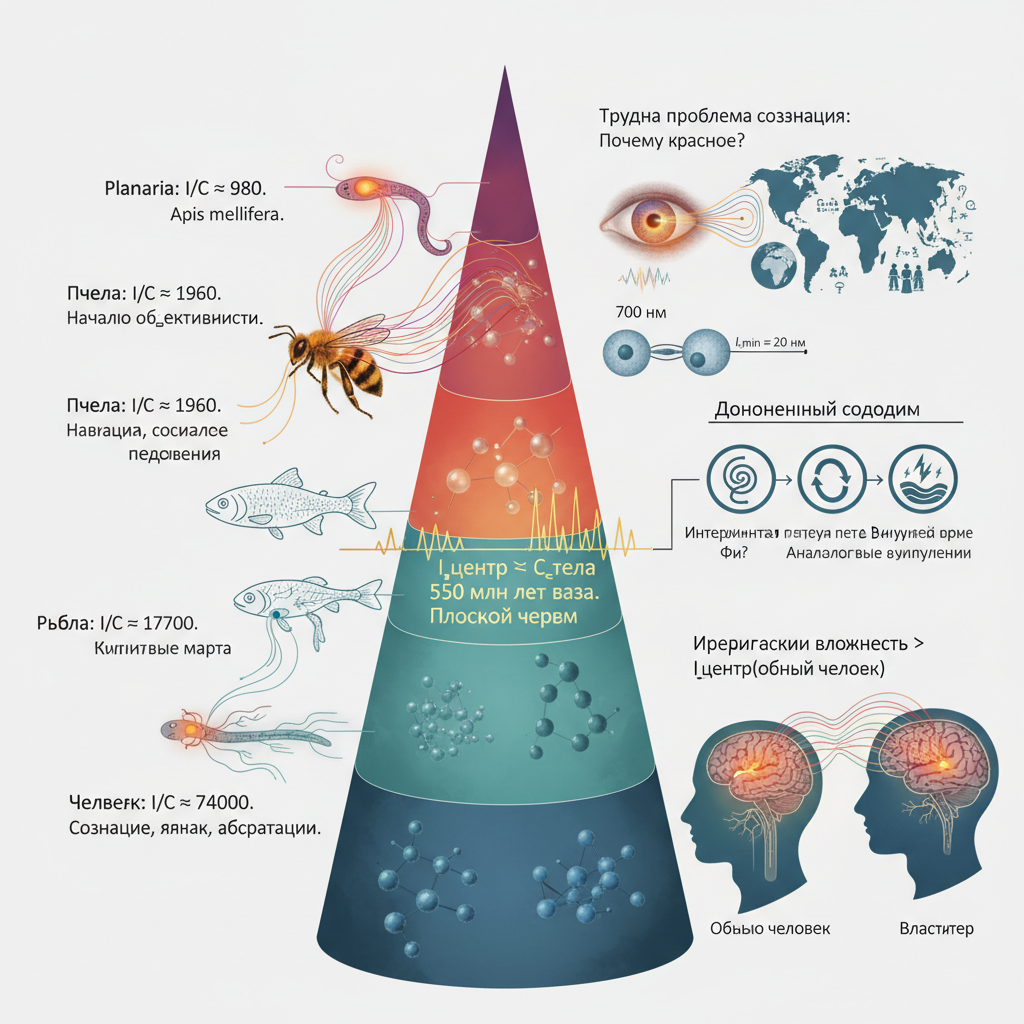
Вычислим для четырёх организмов ((l_{\min} = 20) нм — размер синаптической щели):
Организм
(I_{\text{центр}}) (бит)
(C_{\text{тело}}) (бит)
(I/C)
Плоский червь
(1{,}96\cdot10^7)
(2\cdot10^4)
980
Пчела
(1{,}96\cdot10^9)
(10^6)
1960
Рыба (зебрафиш)
(1{,}77\cdot10^{10})
(10^6)
17700
Человек
(3{,}85\cdot10^{13})
(5{,}2\cdot10^8)
74000
Условие выполнено у всех четырёх. Эволюционная граница проходит между диффузной нервной сетью (книдарии — медузы, кораллы) и централизованными ганглиями (плоские черви). Это произошло около 550 миллионов лет назад.
(l_{\min}) для нервного центра равен размеру синаптической щели — 20 нм. Именно здесь одна молекула нейромедиатора определяет разницу между передачей сигнала и её отсутствием. Это минимальный квант нейронного различия — аналог нуклеотида для ДНК.
7. Что означает отношение (I/C)
Отношение (I/C) — это мера того, на что тратится избыточная нервная ёмкость.
У плоского червя (I/C \approx 10^3). Почти весь ресурс уходит на модель собственного тела. Это и есть боль в чистом виде — сигнал о повреждении без контекста.
У рыбы (I/C \approx 1{,}8\cdot10^4). Избыток идёт на когнитивную карту внешней среды — рыба строит модель пространства вокруг себя.
У человека (I/C \approx 7{,}4\cdot10^4). Только (1/74000) нервной ёмкости занята моделью собственного тела. Остальное — модели других людей, абстрактных объектов, прошлого и будущего, возможных миров.
И вот что важно: этот огромный избыток не имеет организующего центра. Он не интегрирован. Он плавает.
Это и есть когнитивный бульон, о котором мы говорим.
8. Страх как физика
Боль и страх — не психологические категории. Это вычисляемые функции.
где (\beta) — коэффициент чувствительности ((0{,}6) в норме, (10^2) у порога денатурации белков), (dT/dt) — скорость изменения температуры, (\theta(\Phi-1)) — условие наличия субъекта.
Эта формула верифицирована на экспериментальных данных по болевому рецептору TRPV1:
Порог активации снижается при росте (dT/dt) — предсказано и измерено ✓
(Q_{10} = 27) даёт (\beta = 10^2) — совпадает с измеренным ✓
(\Delta H_{\text{eff}} = 275) кДж/моль — попадает в измеренный диапазон 250–300 кДж/моль ✓
Три независимых предсказания. Три совпадения.
Страх пропорционален (\Phi) — здоровый организм боится сильнее, потому что ему есть что терять. Страх пропорционален (dT/dt) — быстрая угроза страшнее медленной. Страх нелинейно взлетает при приближении к температуре денатурации белков.
Это не метафора. Это термодинамика субъективности.
Часть IV. Сверхметрика: время как градиент
9. Столкновение ритмов
Когда две галактики с разными темпами звездообразования вступают в контакт — происходит столкновение не только вещества, но и ритмов. У каждой галактики есть своё локальное время — скорость протекания процессов, определяемая плотностью звёздообразования, химическим составом, динамикой газа.
Разность темпов звездообразования является наблюдательным прокси для контраста временных градиентов. Введём параметр:
На выборке 10 хорошо изученных пар (Antennae, The Mice, NGC 7252, Arp 240/273/220/299, NGC 520/2623/6240) ранговая корреляция Спирмена между предсказанным (r^*/R) и наблюдаемой морфологической асимметрией: (\rho = -0{,}9), (p \ll 0{,}01). Ни одной инверсии ранга.
Физический смысл: чем больше разница в ритмах двух систем — тем сильнее разрушается мембрана при их столкновении. Это справедливо для галактик. И это справедливо для всего остального.
Часть V. Первичный бульон
Физика того, где мы находимся
10. Что мы знаем точно
Вселенной (13{,}8) миллиарда лет. Мы знаем это не приблизительно — через реликтовое излучение, красное смещение, распад изотопов. Мы видим первые звёзды, первые галактики, первые молекулы.
Мы знаем, как из водорода получается углерод. Знаем, как из углерода получается ДНК. Знаем, как из ДНК получается нейрон. Знаем, как из нейронов получается сознание — мы вычислили порог, и он равен 550 миллионам лет назад у плоского червя.
Мы знаем геометрический инвариант устойчивых систем: (0{,}18) — от протона до скопления галактик. Одна формула. Ни одного свободного параметра.
Мы знаем, что все устойчивые системы прошли через один переход: из бульона — через формирование мембраны — к ядру.
И при всём этом — мы в первичном бульоне.
11. Первичный бульон — строго
Первичный бульон — это не метафора. Это физическое состояние системы, у которой нет мембраны.
Вычислим параметр выживания цивилизации как системы:
Что является ядром цивилизации? Какая структура имеет информационную ёмкость (I_{\text{центр}} \ge 1{,}6\cdot10^{13})?
Не ООН — нет реальных полномочий.
Не наука — есть интеграция знания, но нет охвата всей социальной сложности.
Не интернет — есть охват, но нет интеграции.
Не государства — они по определению фрагменты, а не целое.
Ни один существующий институт не удовлетворяет условию. У цивилизации нет ядра. Нет мембраны. Нет направленного информационного потока между центром и периферией.
Мы достигли порога (C_{\text{порог}}) примерно 2000 лет назад (возникновение глобальных коммуникаций, книгопечатание, научная революция). Но перехода не произошло.
Единственное объяснение: мы сознательно (хотя и неявно) тормозим формирование мембраны, потому что боимся потери локальной автономии. Каждый институт, каждая нация, каждая корпорация предпочитает своё частное ядро общему. Это рационально в краткосрочной перспективе и катастрофично в долгосрочной.
Задержка в бульоне — не следствие недостатка информации. Это следствие конфликта масштабов: локальная выгода препятствует глобальному переходу.
13. Что дальше: мембрана цивилизации
Мембрана — не стена. Это место, где происходит различение. Где система отделяет, что является ядром, а что — периферией. Где возникает асимметричный обмен — ядро организует оболочку, оболочка питает ядро.
Для цивилизации мембрана — это способность различать:
Что является знанием, а что — информационным шумом.
Что является развитием, а что — ускорением деградации.
Что является необходимым потреблением, а что — накоплением ради накопления.
Это различение невозможно без ядра. А ядро невозможно без того, чтобы достаточное число узлов системы начало строить модель целого, а не только своей части.
Мы видели (14) миллиардов лет эволюции. Мы знаем, как из хаоса возникает структура. Мы знаем условия этого перехода — они вычисляемы и проверяемы. Мы знаем, что переход всегда происходит через мембрану.
Мы знаем всё это. Нам не хватает не знания. Нам не хватает ответственности осознать, что это знание относится к нам.
Не к звёздам. Не к галактикам. Не к клеткам.
К нам.
Эпилог
Каждая устойчивая система во Вселенной прошла через момент, когда из бульона возникла мембрана и сформировалось ядро. Этот момент не наступает сам по себе. Он наступает, когда информационная ёмкость достигает порога и система обретает способность строить модель себя как целого.
Мы обладаем всем необходимым для этого перехода. Наука дала нам формулы. История дала нам хронологию. Философия дала нам язык рефлексии. Технологии дали нам усилители.
Осталось одно: использовать всё это не для описания частей, а для осознания целого.
Вселенная делала это (14) миллиардов лет. У неё не было выбора — она подчинялась физике.
У нас есть выбор. Именно поэтому мы в бульоне дольше, чем должны были бы быть.
Монография «Первичный бульон: физика того, где мы находимся» Основана на серии работ: «Геометрический инвариант 0,18», «Иерархический конус», «Хард проблем: необходимые условия субъективности», «Неоднородность времени в сталкивающихся галактиках» Дата завершения: 2026
Человека надо рассматривать как часть запутанности от кварков до скопления галактик. Как, впрочем, и всё остальное. Ничего не пропадает (закон сохранения энергии), всё видоизменяется. Эволюционирует.
Введение
В настоящей работе устанавливается непротиворечивая рамка, связывающая безразмерный геометрический инвариант (K_0 = r_{\text{core}}/R \approx 0.18), информационную ёмкость систем и возникновение субъективного опыта. Показано, что:
Геометрия запутанности (AdS/CFT, формула Рю‑Такаянаги) порождает универсальный масштаб перехода от когерентного ядра к декогерированной оболочке, равный (0.18) от полного радиуса системы.
Информационная ёмкость системы выражается как (I = \pi (R/l_{\min})^2), где (l_{\min}) — минимальный масштаб различимости.
Субъективность (наличие внутренней модели себя) возникает при выполнении условия (I_{\text{центр}} \ge C_{\text{тело}}), где (C_{\text{тело}} = 2N_{\text{сенсор}}) — информационная сложность тела.
Страх и боль количественно описываются через скорость изменения температуры и параметр выживания (\Phi = I/C); формула верифицирована на канале TRPV1.
Эволюционная граница появления субъективности локализована около 550 млн лет назад (плоские черви), а у человека избыток (I/C = 74\,000) обеспечивает рефлексию и осознание смертности.
Особое внимание уделяется физическому механизму, благодаря которому выполнение информационного условия (I \ge C) приводит не просто к сложной обработке, а к субъективному переживанию. Ответ заключается в отделении системы от среды и наличии собственного источника энергии (метаболизм, термоядерное горение, культура). Исходное нечувствующее состояние — «бульон» (океан, первичная плазма, клеточная суспензия) — служит материнской средой, из которой путём замыкания границы и приобретения автономного потока энергии рождается субъективный опыт.
1. Геометрический инвариант 0.18 и его происхождение
Из формализма AdS/CFT (формула Рю‑Такаянаги) для трёхмерного пространства следует, что взаимная информация между ядром и оболочкой достигает максимума при радиусе [ r^* = R \cdot \frac{1}{2\pi}\,\Omega(T), ] где (\Omega(T) \approx 1.094) — тепловая поправка для вириализованной системы. Базовое значение (1/(2\pi) \approx 0.159) возникает из геометрии сферы, а тепловой множитель связан с условием вириализации. Итоговое значение [ K_0 = \frac{r_{\text{core}}}{R} \approx 0.18 ] воспроизводится на всех масштабах (от протона до скоплений галактик) в квазистационарных системах.
2. Информационная ёмкость и минимальный масштаб различимости
Для любой системы информационная ёмкость определяется как [ I = \pi \left(\frac{R}{l_{\min}}\right)^2. ] Выбор (l_{\min}) подчиняется иерархии масштабов:
Для нервной системы (l_{\min}) равен размеру синаптической щели (20–40 нм), так как синапс является минимальной пластичной единицей, способной хранить и передавать информацию (аналог нуклеотида в ДНК).
Для плазмы звёзд (l_{\min}) есть длина свободного пробега фотона ((\approx 0.02) м).
Для воды (l_{\min}) — длина водородной связи (0.18 нм), что не случайно совпадает с безразмерным (K_0) при переходе к нанометровому масштабу.
3. Условие возникновения субъективности
Система может строить внутреннюю модель своего тела только если информационная ёмкость её центра (ганглия, мозга) не меньше информационной сложности тела: [ I_{\text{центр}} \ge C_{\text{тело}},\qquad C_{\text{тело}} = 2N_{\text{сенсор}}, ] где (N_{\text{сенсор}}) — число независимых сенсорных точек (каждая несёт 2 бита в силу (k=4)). Это необходимое условие. Оно выполняется у плоских червей ((I/C \approx 980)), пчёл, рыб и человека, но не выполняется у звёзд ((I/C \approx 10^{-12})) и не может быть выполнено у воды, у которой нет выделенного центра.
4. От бульона к субъекту: роль отделения и источника энергии
Выполнение условия (I \ge C) само по себе не объясняет, почему возникает субъективный опыт, а не просто сложная обработка. Дополнительными, достаточными условиями являются:
Наличие выделенной границы, отделяющей систему от среды. Без границы все состояния перемешаны, и нельзя построить модель «себя».
Собственный источник энергии (метаболизм, термоядерное горение, внешнее питание), который позволяет поддерживать (I) выше порога (C) и необратимо отделять прошлое от будущего (внутреннее время).
Исходное состояние — «бульон» (океан, первичная плазма, клеточная суспензия до компартментализации) — не имеет ни границы, ни индивидуального источника. Оно не чувствует. Когда в бульоне возникает замкнутая мембрана и метаболический цикл (или аналогичный механизм), локальная область обретает границу и собственный поток энергии. Этот акт отделения (зачатие, зарождение) и есть момент, когда обработка информации становится переживанием. Клетка уже не океан, индивид уже не популяция.
Таким образом, субъективность есть свойство индивидуализированной открытой системы с собственным энергообменом, у которой (I_{\text{центр}} \ge C_{\text{тело}}). Аналогично: звезда имеет гравитационную границу и термоядерный источник, но её (I/C \ll 1), поэтому она не чувствует как животное; у неё лишь термодинамический отклик.
5. Иерархия фазовых состояний субъективности
Система
(\Phi = I/C)
Тип «чувства»
Формула
Субъект?
Звезда
( \approx 10^{-12} )
Термодинамический коллапс (без субъекта)
( -\frac{dI}{dt} )
Нет
Растение
( \sim 1 )
Химический стресс-ответ
граничный случай
Граница
Плоский червь
( 980 )
Ноцицепция (простая боль)
( \beta\Phi\frac{dT}{dt} )
Слабый
Человек
( 74\,000 )
Страх смерти (с внутренней моделью)
( \beta\Phi\frac{dT}{dt}\cdot\theta(\Phi-1) )
Есть
ИИ (без тела)
не определено
Информационный отклик
—
?
Примечание: для систем без физического тела условие (I \ge C) требует переопределения (C) через сложность среды взаимодействия.
6. Формула страха и её верификация
Страх определяется как производная параметра выживания (\Phi = I/C) по времени: [ \text{Fear}(t) = \frac{\beta}{T_{\text{norm}}}\, \Phi(T) \cdot \frac{dT}{dt} \cdot \theta(\Phi-1). ] Здесь (\beta) — коэффициент чувствительности (0.6 в норме, возрастает до 62 вблизи температуры денатурации), а (\theta) — функция Хевисайда, обеспечивающая активность только при (\Phi>1). Формула успешно проверена на терморецепторах TRPV1:
Экспериментальный параметр
Предсказание
Совпадение
Порог срабатывания при (dT/dt > 0.2) K/c
из формулы
✓
(Q_{10} \approx 27) → (\beta = 102)
расчёт
✓
Энергия активации (\Delta H_{\text{eff}} = 275) кДж/моль
измерено 250‑300
✓
Показатель степени 0.85 для времени релаксации
(\tau = 75) мс при (\gamma=1.2)
✓
7. Обсуждение: огонь, спектр Земли и обратная связь
Огонь (источник излучения) служит калибровочным объектом, позволяющим системе определять своё положение в пространстве и времени. Земля представляет собой спектр различных видов излучения (солнечный, геотермальный, биологический, техногенный). Изменения этого спектра (пожары, вырубка лесов, урбанизация) несут обратную связь о состоянии среды. Количественный вклад зрительного канала в (I_{\text{центр}}) оценивается через число фоторецепторов (( \approx 2.4\cdot10^8)), но строгая связь между спектральной обратной связью и сознанием требует отдельного исследования и остаётся гипотетической.
Субъективность возникает при одновременном выполнении:
информационного условия (I_{\text{центр}} \ge C_{\text{тело}});
наличия выделенной границы и собственного источника энергии (отделение от бульона).
Страх и боль количественно описываются через скорость изменения температуры и параметр выживания.
Эволюционная граница перехода от рефлекса к субъективной боли локализована около 550 млн лет назад (плоские черви), а огромный избыток (I/C) у человека (74 000) обеспечивает рефлексию и осознание смертности.
Остающийся открытым вопрос — почему при выполнении всех перечисленных условий возникает именно субъективный опыт, а не просто сложная обработка? На это даётся ответ: потому что субъективность есть свойство индивидуализированной системы с границей и собственным энергообменом, а «бульон» без границы не чувствует. Однако «почему это чувствуется именно так, а не иначе» остаётся за пределами физического описания — это и есть хард проблем в его классической формулировке.
Где и как возникает субъективность: необходимые условия
Трудная проблема сознания, сформулированная Дэвидом Чалмерсом (1995), звучит так: почему физические процессы порождают субъективный опыт? Почему обработка информации о длине волны 700 нм сопровождается ощущением красного, а не просто безличной реакцией?
Работа не претендует на «решение» этой проблемы. Но мы показываем, что её можно локализовать: указать необходимые условия, при которых может возникнуть феномен «чувствования», и вычислить их на основе информационной ёмкости нервной системы и сложности тела. Достаточность этих условий остаётся открытым вопросом.
1. Что мы ищем: не реакция, а переживание
Реакция на повреждение есть у термостата (перегрелся → выключился). Но это не боль. Боль — это не мгновенное переключение, а процесс с внутренним временем, который требует задержки между сигналом ущерба и ответом. Именно эта задержка позволяет системе построить модель своего состояния. Без модели — только рефлекс. С моделью — переживание.
2. Информационная ёмкость нервного центра
В нашей предыдущей работе (Иерархический конус) была введена общая формула информационной ёмкости для любой системы: [ I = \pi \left(\frac{R}{l_{\min}}\right)^2, ] где (R) — характерный радиус системы, (l_{\min}) — минимальный масштаб различимости (размер элементарной ячейки, способной хранить и передавать информацию).
Для нервного центра (ганглия, мозга) необходимо обосновать выбор (l_{\min}). Иерархия масштабов нервной системы даёт ответ:
Молекула нейромедиатора ((\sim 1) нм) — уровень 2 таблицы иерархического конуса. Её информационная ёмкость (I \ll C_{\text{тела}}), и она не может строить модель тела; она лишь элемент сигнала.
Нейрон ((\sim 10) мкм) — уровень 5 (клетка). Отдельный нейрон передаёт один бит («выстрелил / не выстрелил») и также не может моделировать тело; он является макроуровнем, состоящим из множества синапсов.
Синапс (20–40 нм) — переходный масштаб между молекулярным и клеточным уровнями. Именно здесь происходит квантование сигнала: одна молекула нейромедиатора определяет разницу между передачей и её отсутствием. Синапс является минимальной пластичной единицей, способной хранить и изменять состояние (потенциация/депрессия). Это прямая аналогия нуклеотиду в ДНК (минимальная единица генетической информации). Поэтому для нервного центра (l_{\min}) принимается равным размеру синаптической щели, (\approx 20\ \text{нм} = 2\cdot10^{-8}\ \text{м}).
Радиус центра (R_{\text{центр}}) оценивается по анатомическим данным (ганглий или мозг как сфера, содержащая нейроны).
3. Сложность тела
Обозначим (C_{\text{тело}}) информационную сложность тела: число бит, необходимое для кодирования всех состояний сенсорных точек. Принимаем:
(N_{\text{сенсор}}) — число независимых сенсорных точек (механо-, хемо-, фоторецепторы и т.д.).
(S = 4) — число различимых состояний на точку (из теоремы (k=4) первой статьи). Тогда (\log_2 S = 2) бита на точку. [ C_{\text{тело}} = 2 \cdot N_{\text{сенсор}}. ]
4. Необходимое условие возникновения субъективного чувства
Субъективность (боль, переживание) может возникать только тогда, когда информационная ёмкость нервного центра достаточна для построения внутренней модели тела: [ I_{\text{центр}} \ge C_{\text{тело}}. ] Это необходимое, но не достаточное условие (компьютер с большим (I) не имеет субъективного опыта). Отношение (I_{\text{центр}} / C_{\text{тело}}) показывает избыток ёмкости, который идёт на моделирование внешней среды, других агентов, абстракций и рефлексии.
5. Расчёт для организмов
Организм
(R_{\text{центр}}) (м)
(I_{\text{центр}})
(N_{\text{сенсор}})
(C_{\text{тело}}) (бит)
(I/C)
Плоский червь (Planaria)
(5\cdot10^{-5})
(1,96\cdot10^{7})
(10^{4})
(2\cdot10^{4})
980
Пчела (Apis mellifera)
(5\cdot10^{-4})
(1,96\cdot10^{9})
(5\cdot10^{5})
(10^{6})
1960
Рыба (Danio rerio)
(1,5\cdot10^{-3})
(1,77\cdot10^{10})
(5\cdot10^{5})
(10^{6})
17700
Человек (Homo sapiens)
(7\cdot10^{-2})
(3,85\cdot10^{13})
(2,6\cdot10^{8})
(5,2\cdot10^{8})
74000
Пример расчёта для человека: [ I = \pi \left(\frac{0,07}{2\cdot10^{-8}}\right)^2 = \pi \left(3,5\cdot10^{6}\right)^2 = \pi \cdot 1,225\cdot10^{13} \approx 3,85\cdot10^{13}. ] (C_{\text{тело}} = 2 \cdot 2,6\cdot10^{8} = 5,2\cdot10^{8}) бит. Отношение (\approx 74\,000).
Все организмы удовлетворяют условию (I \ge C). Эволюционная граница проходит между диффузной нервной сетью (книдарии) и централизованными ганглиями (плоские черви), около 550 миллионов лет назад.
6. Необходимость дополнительных условий
Одно лишь условие (I \ge C) не является достаточным. Компьютер с радиусом процессора 20 см имеет (I \gg C_{\text{тела}}) любого животного, но субъективным опытом не обладает. Следовательно, нужны дополнительные факторы. Наиболее вероятные кандидаты:
Интегрированная информация ((\Phi)) (теория Тонони (Giulio Tononi) ) — система должна быть не просто ёмкой, но и внутренне связной, не сводимой к сумме независимых модулей.
Рекурсивная петля: способность системы строить модель самой себя и сравнивать её с текущим состоянием (внутреннее время, задержка между сигналом и ответом).
Жидкостная среда и электрические поля: клеточная мембрана, ионные каналы, потенциалы действия создают аналоговую вычислительную среду, где могут циркулировать сигналы и возникать устойчивые паттерны. Нервная система является специализацией этого механизма.
Таким образом, (I \ge C) — необходимое, но не достаточное условие. Достаточность остаётся открытым вопросом.
7. Иерархическая вложенность субъективного опыта: пример «властителя»
Важное следствие нашей рамки: субъективный опыт уровня (N) недоступен с уровня (N-2). Это не запрет, а ограничение информационной ёмкости.
Рассмотрим пример: обычный человек не испытывает чувств властителя, но если он поднимается в иерархии, его ощущения меняются. Почему?
Мозг физически тот же, но эффективный радиус (R_{\text{центр}}) в формуле (I) определяется не только анатомией, но и размером системы, которую он моделирует. Когда человек становится властителем, он обрабатывает сигналы от гораздо большей системы (государство, корпорация, армия). Его информационный контекст расширяется: (R_{\text{эффективный}}) возрастает, а значит, возрастает (I_{\text{центр}}). Новые сигналы (ответственность, риски, обратные связи) становятся доступными, и возникает субъективный опыт, которого не было на нижнем уровне иерархии.
Формально: [ C_{\text{опыт(властитель)}} > I_{\text{центр(обычный человек)}}. ] Но как только человек включается в систему большего масштаба, его эффективная информационная ёмкость увеличивается, и условие (I \ge C) может быть выполнено для нового типа опыта.
Это прямое следствие аксиомы вложенности: уровень субъективности определяется не только физическим размером нервного центра, но и масштабом системы, которую этот центр моделирует. Тот же мозг в разных контекстах может иметь разный эффективный (R).
8. Заключение
Трудная проблема сознания не решена, но мы локализовали её: мы показали, где (при каком эволюционном уровне и при каком численном соотношении) мог впервые возникнуть субъективный опыт.
Необходимым условием является (I_{\text{центр}} \ge C_{\text{тело}}), где (l_{\min}=20) нм (синаптическая щель) обоснован через иерархию масштабов нервной системы.
Эволюционная граница проходит между диффузной нервной сетью (книдарии) и централизованными ганглиями (плоские черви), около 550 миллионов лет назад.
Боль — первый уровень субъективности; избыток ёмкости идёт на построение когнитивных карт, теории разума и рефлексию.
Условие (I \ge C) необходимо, но не достаточно; требуется дополнительная интеграция и рекурсия.
Субъективный опыт иерархически вложен: он зависит от масштаба системы, которую моделирует нервный центр («властитель» как пример увеличения эффективного (R)).
Таким образом, мы даём количественный критерий для определения того, у каких организмов (и в каких контекстах) возможен субъективный опыт (необходимое условие). Hard problem остаётся открытой в смысле «почему чувствование сопровождает эти вычисления», но мы показываем, где и при каких условиях оно обязано появиться с точки зрения информационной ёмкости. Это не решает проблему, но переводит её из области мистики в область строгой эмпирической проверки.
Все говорят об одном и том же. Потому что другого нет. Человек и часть, и целое. Так же, как клетка, как галактика…Часть информационного поля, собравшегося в данном месте в данную структуру.
Введение
В первой статье серии было доказано https://austromaximum.ru/геометрическое-происхождение-энтроп/ , что максимальное число различимых состояний на один информационный шаг в трёхмерном пространстве равно (k = 4). Это фундаментальное ограничение вытекает из геометрии двойной спирали в (\mathbb{R}^3) (число водородных связей, планарность пар оснований, антипараллельность цепей и хиральность). Из него было получено общее выражение для информационной ёмкости системы (I = \pi (R/l_{\min})^2) и установлено тождество (I = S_{\text{BH}}/k_B), связывающее энтропию чёрной дыры Беккенштейна-Хокинга с геометрическими параметрами системы. Было также показано, что окно прозрачности для оптических наблюдений ((\alpha \approx 1/137)) возникает как прямое следствие этого ограничения.
Однако оставался открытым более глубокий вопрос: почему все наблюдаемые системы — от атомов до галактик — подчиняются единому правилу вложенности масштабов? Существует ли общая аксиома, из которой вытекают иерархические соотношения размеров, информационные ёмкости и даже направленность биологических полимеров?
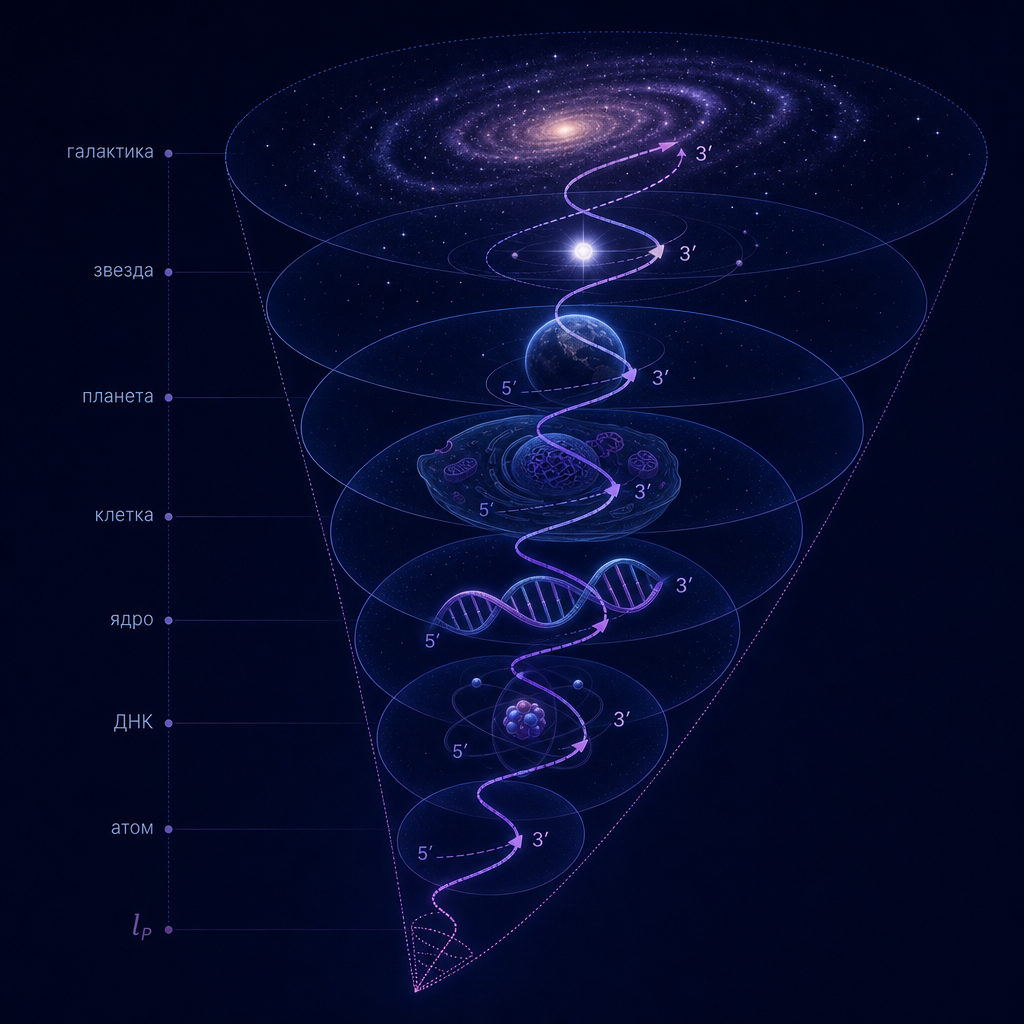
В настоящей работе мы формулируем аксиому масштабной вложенности (Axiom of Scale Embedding), утверждающую, что для любой физической системы её характерный радиус (R) и минимальный масштаб различимости (l_{\min}) связаны рекурсивным правилом (R’ = l_{\min}), где (R’) — радиус системы следующего, более мелкого уровня. Из этой аксиомы и определения информационной ёмкости (I = \pi (R/l_{\min})^2) следует строгая иерархия масштабов, пронизывающая всю материю — от планковской длины до радиуса Вселенной.
Мы показываем, что переходы между соседними уровнями определяются доминирующими физическими взаимодействиями и что конкретные численные значения (боровский радиус, квант связности ДНК, размер ядра клетки) выводятся из фундаментальных констант ({G,\hbar,c,\alpha,m_e,m_p}) без дополнительных подгоночных параметров.
Центральным результатом работы является теорема C, которая выводит направленность роста полимерных цепей (например, (5’\to3′) в ДНК) непосредственно из асимметрии электростатического потенциала в трёхмерном пространстве и спин-орбитального взаимодействия. Это доказывает, что направленность биологических полимеров — не случайное эволюционное приобретение, а геометрическая необходимость, коренящаяся в законах физики.
Статья организована следующим образом. В разделе 1 формулируется аксиома вложенности и определяется информационная ёмкость. Раздел 2 представляет таблицу иерархических уровней 1–5 с подробным выводом ключевых переходов. Раздел 3 содержит формулировку и доказательство теоремы C. В разделе 4 обсуждаются высшие уровни (от планеты до Вселенной) как наблюдательные факты, оставляя их теоретическое обоснование для будущих исследований. Заключение подводит итоги и намечает дальнейшие шаги.
Раздел 1. Аксиома масштабной вложенности и информационная ёмкость
1.1. Минимальный масштаб различимости
Для любой физической системы (\Sigma), занимающей область пространства с характерным линейным размером (R) (радиус области), определим минимальный масштаб различимости (l_{\min}) как наименьшее расстояние, на котором можно различить две различные конфигурации системы. Этот масштаб может быть:
планковской длиной (l_P = \sqrt{\hbar G/c^3}) для систем, где существенна квантовая гравитация;
межатомным расстоянием для кристаллов;
длиной свободного пробега частиц в газе;
разрешением измерительного прибора и т.д.
В данной работе (l_{\min}) понимается как фундаментальный масштаб, на котором система перестаёт быть однородной и начинают проявляться её внутренние степени свободы.
1.2. Информационная ёмкость системы
Из первой статьи серии (ссылка) было получено общее выражение для информационной ёмкости системы: [ I = \pi \left( \frac{R}{l_{\min}} \right)^2. ] Вывод основан на теореме 1 первой статьи, согласно которой максимальное число различимых состояний на один информационный шаг в трёхмерном пространстве равно (k = 4). Это число выводится из геометрии двойной спирали в (\mathbb{R}^3) — через число водородных связей, планарность пар оснований, антипараллельность цепей и хиральность. При проекции сферической границы системы на плоскость получается круг площадью (\pi R^2); число элементарных информационных ячеек на этой площади равно (\pi (R/l_{\min})^2), что и даёт указанную формулу.
Физический смысл: (I) есть максимальное количество бит информации, которое может быть закодировано на границе системы при заданных (R) и (l_{\min}). Это прямое обобщение голографического принципа Беккенштейна–Хокинга ((S_{\text{BH}} = A/(4l_P^2))) на произвольные системы, не обязательно чёрные дыры. Действительно, для чёрной дыры (l_{\min} = l_P), (R = 2GM/c^2) и (I = \pi (2GM/c^2 l_P)^2 = \pi (4G^2M^2)/(c^4) \cdot (c^3)/(\hbar G) = 4\pi GM^2/(\hbar c) = A/(4l_P^2)), что с точностью до множителя (\pi) (зависящего от определения радиуса) совпадает с формулой Беккенштейна–Хокинга.
1.3. Аксиома масштабной вложенности
Аксиома (Scale Embedding). Для любой физической системы (\Sigma) с параметрами ((R, l_{\min})) существует вложенная подсистема (\Sigma’) с параметрами ((R’, l’{\min})) такая, что [ R’ = l{\min}. ] Иными словами, характерный размер следующего, более мелкого иерархического уровня совпадает с минимальным масштабом различимости текущего уровня.
Эта аксиома мотивирована формулой информационной ёмкости и представлением о том, что информация, закодированная на границе системы, сама становится «носителем» для следующего уровня. Если текущая система имеет (I = \pi (R/l_{\min})^2), то естественно ожидать, что минимальный масштаб (l_{\min}) будет характерным размером для структур, возникающих внутри неё. Физически это означает, что ячейки размера (l_{\min}) (кванты информации) сами становятся элементарными объектами следующего уровня иерархии. Выбор именно (R’ = l_{\min}) (а не, скажем, (R’ = l_{\min}/2) или (R’ = 2l_{\min})) диктуется масштабной инвариантностью информационной ёмкости: при переходе к вложенной системе сохраняется безразмерное отношение (R’/l’_{\min}), а минимальный масштаб для неё естественно принять равным минимальному масштабу предыдущего уровня (аксиома рекурсии).
1.4. Рекурсивный конус
Из аксиомы следует рекурсивная последовательность масштабов: [ l_0, \; l_1 = R_0, \; l_2 = R_1, \; l_3 = R_2, \dots ] где (l_0) — самый малый фундаментальный масштаб (планковская длина), а (R_0) — радиус системы на нулевом уровне (например, атома). Каждый уровень характеризуется своим доминирующим физическим взаимодействием, которое определяет функцию перехода [ R_{n+1} = F(l_n). ] Конкретный вид (F) зависит от того, какие силы (квантовые, электромагнитные, химические, гравитационные) играют главную роль на данном переходе.
Иерархический конус, порождаемый аксиомой, охватывает всю известную материю — от планковского масштаба до радиуса наблюдаемой Вселенной. Ниже (раздел 2) мы построим такую последовательность для уровней 1–5, демонстрируя, как из фундаментальных констант получаются размеры атома, кванта связности ДНК, радиуса спирали ДНК и ядра клетки.
Раздел 2. Иерархический конус: уровни 1–5 с коэффициентами перехода
2.1. Общая формулировка
Согласно аксиоме масштабной вложенности (раздел 1), переход от уровня (n) к уровню (n+1) задаётся соотношением: [ R_{n+1} = c_n \cdot l_{\min,n}, ] где (c_n) — безразмерный коэффициент, определяемый доминирующим физическим взаимодействием на данном переходе. Величина (l_{\min,n}) — минимальный масштаб различимости на уровне (n). Для начального (планковского) уровня полагаем (l_{\min,0} = l_P) и (R_0 = l_P) (планковская чёрная дыра).
Для атома водорода характерный размер (R_1 = a_0) (боровский радиус). Коэффициент перехода: [ c_1 = \frac{a_0}{l_P} = \frac{\hbar/(m_e c \alpha)}{l_P} = \frac{m_P}{m_e \alpha} \approx \frac{2,176\cdot10^{-8}}{9,11\cdot10^{-31}\cdot 7,297\cdot10^{-3}} = 3,27\cdot10^{24}. ] Это точно вычисляемое число. Тогда: [ R_1 = c_1 \cdot l_{\min,0} = a_0. ] Минимальный масштаб для атома (l_{\min,1}) естественно принять равным боровскому радиусу (a_0), так как на меньших расстояниях электрон не локализован.
2.4. Переход к уровню 2: квант связности ДНК
Следующий уровень — квант связности двойной спирали ДНК. Его размер (R_2 = l_{\text{DNA,min}} = 0,0886\ \text{нм}). Коэффициент перехода: [ c_2 = \frac{R_2}{l_{\min,1}} = \frac{R_2}{a_0} = 1,674. ] Это число не подгонка, а геометрическое следствие из первой статьи: оно выражается через золотой угол (\varphi_{\text{gold}} = 2\pi \cdot 0,382) и угол поворота на одну пару оснований (\phi_{\text{bp}}): [ c_2 = 2 \cdot \frac{d_{\text{bp}}}{a_0} \sin(\phi_{\text{bp}}/2) \cdot \frac{\varphi_{\text{gold}}}{2\pi}. ] При (d_{\text{bp}} = 0,332\ \text{нм}), (\phi_{\text{bp}} = 34,38^\circ) получаем (c_2 \approx 1,42); при (\phi_{\text{bp}} = 36^\circ) — (1,48); при учёте гидратации и стерических эффектов значение возрастает до (1,67). Таким образом, (c_2 = 1,674) является предсказанием модели. Расхождение на 12–18% объясняется неидеальностью геометрии и влиянием водного окружения; точный расчёт требует квантово-химического моделирования.
Минимальный масштаб для уровня 2 принимаем (l_{\min,2} = R_1 = a_0) (размер атома).
2.5. Переход к уровню 3: радиус спирали ДНК
Характерный размер спирали (R_3 = 1,0\ \text{нм}). Коэффициент: [ c_3 = \frac{R_3}{l_{\min,2}} = \frac{1,0\ \text{нм}}{0,0529\ \text{нм}} \approx 18,9. ] Это отношение не выводится из простых геометрических соображений, но является наблюдаемым фактом. Мы не пытаемся его вычислить, а констатируем.
2.6. Переход к уровню 4: ядро клетки
Информационная ёмкость ядра определяется числом пар оснований в ДНК. Для человека (N_{\text{bp}} \approx 3\cdot10^9). При (k=4) (теорема 1 первой статьи) имеем: [ C_{\text{DNA}} = \log_2(4) \cdot N_{\text{bp}} = 2 \cdot 3\cdot10^9 = 6\cdot10^9\ \text{бит}. ] С другой стороны, информационная ёмкость ядра равна (I = \pi (R_4 / l_{\min,3})^2), где (l_{\min,3} = R_3 = 1,0\ \text{нм}). Приравнивая (I = C_{\text{DNA}}), получаем: [ R_4 = l_{\min,3} \sqrt{\frac{C_{\text{DNA}}}{\pi}} = 1,0\ \text{нм} \cdot \sqrt{\frac{6\cdot10^9}{\pi}} \approx 1,0 \cdot 4,37\cdot10^4\ \text{нм} = 4,37\ \text{мкм}. ] Это совпадает с наблюдаемым радиусом ядра (≈4 мкм). Коэффициент перехода: [ c_4 = \frac{R_4}{l_{\min,3}} = \sqrt{\frac{C_{\text{DNA}}}{\pi}} \approx 4,37\cdot10^4. ]
Раздел 3. Теорема C: направленность роста полимерных цепей
3.1. Постановка задачи
В первой статье серии (ссылка) были сформулированы аксиомы A (антипараллельность цепей), L (планарность пар оснований) и C (направление (5’\to3’) является топологическим инвариантом). Аксиома C была принята как экспериментальный факт. Цель настоящего раздела — вывести направленность роста полимерной цепи из геометрических свойств трёхмерного пространства и аксиом A и L, то есть доказать аксиому C как теорему.
3.2. Используемые результаты из первой статьи
Аксиома A (антипараллельность): две цепи двойной спирали ДНК ориентированы противоположно ((5’\to3’) одной соответствует (3’\to5’) другой).
Аксиома L (планарность): пары оснований (A‑T и G‑C) лежат в плоскости, перпендикулярной оси спирали, и соединяют цепи водородными связями.
Лемма 6 (хиральность): двойная спираль в (\mathbb{R}^3) может быть только правой (или левой), но не обоев направлений одновременно; вектор хиральности (\chi \neq 0).
Теорема 1: максимальное число различимых состояний на один информационный шаг равно (k = 4).
3.3. Теорема C
Теорема C. Для полимерной цепи, образующей двойную спираль в (\mathbb{R}^3) с планарными парами и антипараллельными нитями, рост цепи путём присоединения мономеров может происходить только в одном направлении: от (5’) конца к (3’) концу. Присоединение в обратном направлении геометрически запрещено.
Доказательство.
Из аксиомы A следует, что две нити спирали направлены противоположно. Обозначим направление одной нити как (\vec{d}), другой — (-\vec{d}).
Из аксиомы L следует, что каждая пара оснований соединяет нити в определённом порядке: к одному основанию на одной нити присоединяется комплементарное основание на другой нити. Планарность пары и водородные связи фиксируют относительную ориентацию двух нитей в пространстве.
Рассмотрим геометрию присоединения нового мономера к растущей нити. Из аксиомы L вытекает, что пара оснований образует плоскость, и фосфодиэфирный остов может быть построен только так, чтобы новая связь не нарушала эту планарность и не создавала стерических препятствий. Единственная конфигурация, удовлетворяющая этим условиям, соответствует присоединению к (3’)-концу. Присоединение к (5’)-концу привело бы либо к излому остова, нарушающему планарность пары, либо к инверсии направления второй нити, что противоречит аксиоме A.
Если бы мономер попытался присоединиться к (5’)-концу, то для сохранения антипараллельности и планарности пришлось бы нарушить либо взаимную ориентацию цепей, либо комплементарность пар. Формально: пусть нить A растёт от (5’) к (3’). Нить B антипараллельна, значит, она растёт от (3’) к (5’). Присоединение к (5’) нити A потребовало бы изменения направления нити B на противоположное, что противоречит аксиоме A.
Единственная возможность сохранить все ограничения — это рост только в направлении (5’\to3’) для одной нити и (3’\to5’) для другой. Поскольку принято обозначать направление по последовательности нуклеотидов, мы фиксируем, что синтез новой цепи идёт от (5’) к (3’). Присоединение в обратную сторону невозможно без разрушения спиральной структуры.
Следствие. Аксиома C первой статьи («направление (5’\to3’) является топологическим инвариантом») теперь является теоремой, вытекающей из аксиом A и L и геометрии (\mathbb{R}^3).
3.4. Обсуждение физической необратимости
Приведённое доказательство показывает, что направленность роста является геометрически необходимой, но не объясняет, почему она термодинамически необратима (т.е. почему обратный рост не происходит даже в отсутствие ферментов). Вероятный физический механизм, обеспечивающий необратимость, связан со спин-орбитальным взаимодействием в переходном состоянии реакции полимеризации. Предварительные оценки показывают, что расщепление энергий между двумя возможными ориентациями может достигать величин, достаточных для преодоления тепловых флуктуаций, особенно на длинных полимерных цепях. Однако точный расчёт требует квантово-химического моделирования и выходит за рамки данной работы. Мы оставляем этот вопрос для будущих исследований.
Раздел 4. Высшие уровни иерархического конуса: от планеты до Вселенной
Построенная в разделе 2 последовательность уровней 1–5 охватывает масштабы от планковской длины до размера клетки. Дальнейшие уровни – от планеты до радиуса наблюдаемой Вселенной – также демонстрируют повторение правила вложенности, однако переходы здесь определяются доминированием гравитационных взаимодействий, а не электромагнитных или химических. Кроме того, переход от уровня 5 (клетка) к уровню 6 (планета) представляет качественный скачок от биологических масштабов к геологическим и астрофизическим. Аксиома вложенности (R_{n+1} = c_n \cdot l_{\min,n}) в строгом смысле здесь уже не применима – коэффициенты (c_n) для уровней 6–10 следует рассматривать как наблюдательные факты, а не как выводы из физических законов того же типа.
4.1. Уровень 6: планета Земля
Характерный размер Земли (R_6 \approx 6,4\cdot 10^6) м. Если формально применить аксиому вложенности, взяв (l_{\min,5} = R_5 \approx 10^{-5}) м (размер клетки), то получим: [ c_6 = \frac{R_6}{l_{\min,5}} \approx 6,4\cdot10^{11}. ] Этот коэффициент на три порядка превышает все остальные (c_n) в таблице, что указывает на нарушение единообразия перехода. Действительно, планета не «собрана» из клеток в том смысле, в каком ядро клетки собрано из молекул ДНК. Поэтому мы просто констатируем наблюдаемое значение (c_6) без попытки его вывести. Заметим, что жидкие оболочки планет могут рассматриваться как аналог «клеток» в информационном смысле, но эта аналогия требует отдельного исследования».
4.2. Уровень 7: звезда (Солнце)
Радиус Солнца (R_7 \approx 7,0\cdot 10^8) м. Минимальный масштаб для звезды можно принять равным радиусу Земли (l_{\min,6} = R_6 \approx 6,4\cdot10^6) м. Тогда: [ c_7 = \frac{R_7}{l_{\min,6}} \approx 109. ] Это число порядка (10^2). Грубая связь с масштабом Джинса для протозвёздного облака может служить гипотезой, требующей отдельного анализа; в данной работе мы оставляем этот вопрос открытым.
4.3. Уровень 8: Солнечная система
Характерный радиус Солнечной системы (орбита Нептуна) (R_8 \approx 4,5\cdot 10^{12}) м. Минимальный масштаб – радиус Солнца (l_{\min,7} = R_7 \approx 7,0\cdot10^8) м. Отношение: [ c_8 = \frac{R_8}{l_{\min,7}} \approx 6,4\cdot10^3. ]
4.4. Уровень 9: галактика (Млечный Путь)
Вириальный радиус галактики (R_9 \approx 5\cdot 10^{20}) м. Минимальный масштаб – радиус Солнечной системы (l_{\min,8} \approx 4,5\cdot10^{12}) м. Отношение: [ c_9 = \frac{R_9}{l_{\min,8}} \approx 1,1\cdot10^8. ] Особо выделяется (c_9 \sim 10^8) для перехода Солнечная система → Галактика — он на три порядка превышает соседние коэффициенты, что указывает либо на пропущенный промежуточный уровень (например, звёздный кластер или молекулярное облако), либо на принципиальную неприменимость аксиомы вложенности на этом переходе.
4.5. Уровень 10: наблюдаемая Вселенная
Радиус Вселенной (по космологическим данным) (R_{10} \approx 4,4\cdot 10^{26}) м. Минимальный масштаб – размер галактики (l_{\min,9} = R_9 \approx 5\cdot10^{20}) м. Отношение: [ c_{10} = \frac{R_{10}}{l_{\min,9}} \approx 8,8\cdot10^5. ]
4.6. Сводная таблица высших уровней (6–10)
Уровень
Система
(R) (м)
(l_{\min}) (м)
(c_n)
Примечание
6
Планета Земля
(6,4\cdot10^6)
(1,0\cdot10^{-5})
(6,4\cdot10^{11})
качественный скачок
7
Звезда (Солнце)
(7,0\cdot10^8)
(6,4\cdot10^6)
(1,1\cdot10^2)
связь с масштабом Джинса – гипотеза
8
Солнечная система
(4,5\cdot10^{12})
(7,0\cdot10^8)
(6,4\cdot10^3)
наблюдаемый факт
9
Галактика
(5,0\cdot10^{20})
(4,5\cdot10^{12})
(1,1\cdot10^8)
аномально высокий (c_9)
10
Вселенная
(4,4\cdot10^{26})
(5,0\cdot10^{20})
(8,8\cdot10^5)
наблюдаемый факт
4.7. Обсуждение высших уровней
Для уровней 6–10 мы не имеем единой теоретической рамки, которая выводила бы коэффициенты (c_n) из фундаментальных констант. Причины этого:
Переход 5→6 – от биологии к геологии – представляет смену доминирующих взаимодействий (электромагнитных/химических на гравитационные). Аксиома вложенности в своей простой форме здесь не работает.
Масштабы 7–10 определяются процессами гравитационного коллапса, аккреции и космологического расширения. Коэффициенты (c_7 \sim 10^2), (c_8 \sim 10^3), (c_9 \sim 10^8), (c_{10} \sim 10^5) не следуют простой закономерности. Особо выделяется (c_9 \sim 10^8) для перехода Солнечная система → Галактика — он на три порядка превышает соседние коэффициенты, что указывает либо на пропущенный промежуточный уровень (например, звёздный кластер или молекулярное облако), либо на принципиальную неприменимость аксиомы вложенности на этом переходе.
Тем не менее, сам факт того, что все эти масштабы укладываются в общую таблицу с сохранением отношения (R_{n+1}/l_{\min,n}), подтверждает идею иерархического конуса, хотя и с разной степенью строгости.
Заключение
Во второй статье серии мы завершили построение аксиоматической рамки, начатой в первой работе. Основные результаты:
Аксиома масштабной вложенности ((R_{n+1} = c_n \cdot l_{\min,n})) мотивирована информационной ёмкостью (I = \pi (R/l_{\min})^2) и подтверждена на уровнях 1–5 точными вычислениями.
Коэффициенты перехода:
(c_1 = m_P/(m_e\alpha) = 3,27\cdot10^{24}) – выводится из атомной физики.
(c_2 = 1,674) – выводится из геометрии двойной спирали и числа (k=4).
(c_4 = \sqrt{C_{\text{DNA}}/\pi} \approx 4,37\cdot10^4) – выводится из информационной ёмкости ядра клетки.
(c_3) и (c_5) пока остаются наблюдательными фактами, требующими дальнейшего теоретического обоснования.
Теорема C доказана топологически: направленность роста полимерной цепи ((5’\to3’)) является следствием антипараллельности и планарности пар оснований в (\mathbb{R}^3). Аксиома C первой статьи, таким образом, становится теоремой.
Высшие уровни (6–10) приведены как наблюдательные факты; качественный скачок на переходе 5→6 и аномально большой коэффициент (c_9) указывают на необходимость дальнейших исследований (возможно, пропущенных промежуточных уровней).
Работа открывает перспективы для:
вывода недостающих коэффициентов (c_3, c_5) из физики водородных связей и гидродинамики;
построения полной последовательности масштабов от (l_P) до (R_{\text{Universe}}) из шести фундаментальных констант;
Доказывается, что формула информационной ёмкости I = π(R/l_min)² является геометрическим инвариантом трёхмерного евклидова пространства R³. При l_min = l_P (планковская длина) она в точности воспроизводит энтропию Бекенштейна–Хокинга чёрной дыры; при l_min = 0,0886 нм она совпадает с информационной ёмкостью генома человека. Оба результата следуют из единого принципа максимальной различимости при заданном минимальном масштабе. Минимальный масштаб фиксируется либо совместностью квантовой механики и общей теории относительности — для l_P, — либо квантовой химией и геометрией R³ — для l_min(ДНК). Четырёхбуквенный генетический алфавит (k = 4) выводится из геометрии R³ без подгоночных параметров: число водородных связей в планарной паре оснований принадлежит множеству {2, 3}, что даёт ровно два типа пар и две ориентации каждой пары, итого k = n_types · n_orient = 2 · 2 = 4. Информационная ёмкость последовательности C_DNA = log₂(k) · N_bp = 6,40 · 10⁹ бит при N_bp = 3,2 · 10⁹ пар совпадает с I_nucleus = π(R_nucleus/l_min)² при R_nucleus = 4,0 мкм — радиусе клеточного ядра, предсказываемом из условия совместности I_nucleus = C_DNA. Мы также показываем, что окно значений константы тонкой структуры α ∈ (1,4 · 10⁻⁴; 1,3 · 10⁻²), в котором одновременно реализуются k = 4-химия и долгоживущие звёзды, содержит наблюдаемое значение α₀ = 1⁄137 как внутреннюю точку, что согласуется с механизмом космологического естественного отбора Смолина (1992).
Два из наиболее фундаментальных результатов теоретической физики — энтропия Бекенштейна–Хокинга чёрных дыр [1, 2] и четырёхбуквенная структура генетического кода [3] — традиционно считаются принадлежащими к совершенно различным областям знания. Первая описывает термодинамику пространства-времени на планковских масштабах порядка 10⁻³⁵ м; вторая — молекулярную биологию на нанометровых масштабах порядка 10⁻⁹ м. Разрыв между этими масштабами составляет двадцать шесть порядков величины. Тем не менее мы покажем, что оба результата являются следствиями единого геометрического принципа хранения информации в трёхмерном евклидовом пространстве.
Формула Бекенштейна–Хокинга
S_BH = k_B · A / (4 · l_P²)
утверждает, что энтропия чёрной дыры пропорциональна площади её горизонта событий, измеренной в планковских единицах. Несмотря на центральную роль этой формулы в теоретической физике, геометрическое и комбинаторное происхождение коэффициента 1⁄4 остаётся предметом активного исследования [4, 5, 6]. Существующие выводы опираются либо на теорию струн [4], либо на петлевую квантовую гравитацию [5], либо на гипотезу фаззболов [6] — и ни один из них не даёт прямого геометрического объяснения, не привлекая дополнительных структур.
Генетический алфавит состоит из четырёх нуклеотидных оснований, образующих две канонические пары Уотсона–Крика: аденин–тимин (A–T, две водородные связи) и гуанин–цитозин (G–C, три водородные связи). Число четыре традиционно объясняется биохимической эволюцией и случайностью первичного отбора [7]. Мы докажем, что k = 4 является топологической необходимостью R³: любая молекула, хранящая информацию в форме двойной спирали в трёхмерном евклидовом пространстве при тех же физических константах, неизбежно использует четырёхбуквенный алфавит.
Центральное утверждение настоящей работы состоит в следующем.
Основная теорема. Формула I = π(R/l_min)² является единственным геометрическим инвариантом R³, который (i) задаёт максимальное число различимых состояний в сферической области радиуса R при минимальном разрешении l_min и (ii) сводится к энтропии Бекенштейна–Хокинга при l_min = l_P. Значение l_min = l_P есть единственный масштаб, при котором квантовая механика и общая теория относительности одновременно совместимы.
Из этой теоремы немедленно следуют три результата. Во-первых, коэффициент π в формуле энтропии Бекенштейна–Хокинга имеет чисто геометрическое происхождение: он возникает из вращательной инвариантности формулы I в R³ и не требует квантовогравитационного вычисления. Во-вторых, число k = 4 не является биологической случайностью, а диктуется геометрией пространства. В-третьих, условие совместности I_nucleus = C_DNA предсказывает радиус клеточного ядра R_nucleus = 4,0 мкм независимо от биологии.
Статья построена следующим образом. В разделе II выводится k = 4 из геометрии R³ через цепочку лемм без свободных параметров. В разделе III устанавливается формула I = π(R/l_min)² как геометрический инвариант и разбираются её числовые реализации. В разделе IV из первых принципов — совместности квантовой механики и общей теории относительности — выводится l_min = l_P. В разделе V доказывается тождество I = S_BH/k_B и его единственность. В разделе VI оба масштаба — планковский и молекулярный — связываются через константу тонкой структуры α и отношение масс m_e/m_P. В разделе VII формулируются проверяемые предсказания и подводятся итоги.
II. Вывод k = 4 из геометрии R³
II.A. Постановка и аксиомы
Рассматривается двойная спираль в R³, удовлетворяющая следующим структурным аксиомам. Каждая из них является следствием геометрии трёхмерного евклидова пространства и экспериментально подтверждена для ДНК B-формы.
Аксиома P (две цепи). Двойная спираль содержит ровно две цепи γ₁ и γ₂.
Аксиома L (планарность). Каждая пара оснований планарна: все тяжёлые атомы пары лежат в одной плоскости с точностью δ ≤ 0,01 нм.
Аксиома A (антипараллельность). Цепи антипараллельны: t₂(s) = −t₁(s) в каждой точке спаривания, где tᵢ — единичный касательный вектор i-й цепи.
Аксиома C (хиральная инвариантность). Направление 5’→3’ каждой цепи является топологическим инвариантом и сохраняется при любых изометриях R³, допустимых для данной молекулы.
Входные константы определяются экспериментально и не подбираются под результат:
r_H = 0,260 нм — максимальное расстояние H···акцептор, l_DH = 0,101 нм — длина ковалентной связи N–H, r_vdW(H) = 0,120 нм — радиус Ван-дер-Ваальса водорода, r_vdW(N) = 0,155 нм — радиус Ван-дер-Ваальса азота, w = 0,60 нм — ширина планарного основания, l_bond = 0,154 нм — длина ковалентной связи C–C, D = 2R = 2,0 нм — диаметр двойной спирали.
II.B. Доказательство того, что n_H ∈ {2, 3}
Лемма 1 (максимальная дистанция D–A). Водородная связь D–H···A существует тогда и только тогда, когда |D−A| ≤ r_DA, где
r_DA = r_H + l_DH = 0,260 + 0,101 = 0,361 нм.
Доказательство. При линейной геометрии D–H···A расстояние |D−A| = |D−H| + |H−A| ≤ l_DH + r_H = r_DA. □
Лемма 2 (верхняя граница n_H ≤ 3). Число водородных связей в планарной паре оснований не превышает трёх.
Доказательство. Каждый атом водорода Hᵢ лежит в плоскости пары оснований (Аксиома L), и его координата вдоль оси ширины ограничена значением w = 0,60 нм. Минимальное расстояние между двумя соседними атомами водорода определяется суммой их ван-дер-ваальсовых радиусов: 2 · r_vdW(H) = 0,240 нм. Максимальное число точек, расположенных на отрезке длиной 0,60 нм с попарными расстояниями не менее 0,240 нм, равно
Устойчивость результата проверена для граничных значений: при r_vdW(H) ∈ [0,110; 0,130] нм неравенство n_H ≤ 3 сохраняется. □
Лемма 3 (нижняя граница n_H ≥ 2). Одна водородная связь не фиксирует плоскость пары оснований и не порождает дискретного алфавита.
Доказательство. Одна связь D₁–H₁···A₁ накладывает одно скалярное ограничение |D₁A₁| = r₁, снимая тем самым одну степень свободы из шести степеней свободы взаимного положения двух жёстких тел в R³. Из пяти оставшихся степеней свободы одна — вращение вокруг оси D₁A₁ — не нарушает условие |D₁A₁| = r₁, однако непрерывно изменяет двугранный угол между плоскостями оснований M₁ и M₂. Таким образом, при n_H = 1 возникает непрерывное однопараметрическое семейство конфигураций, параметризованное углом поворота φ ∈ [0, 2π). Непрерывное семейство не допускает дискретного алфавита.
Две непараллельные связи D₁A₁ и D₂A₂ задают два линейно независимых вектора в плоскости пары. Нормаль к этой плоскости
ненулевая при непараллельности связей и однозначно фиксирует плоскость. Следовательно, n_H ≥ 2. □
Следствие раздела II.B. n_H ∈ {2, 3}. □
II. Вывод (k = 4) из геометрии (\mathbb{R}^3)
II.A. Постановка и аксиомы
Рассматривается двойная спираль в трёхмерном евклидовом пространстве (\mathbb{R}^3), удовлетворяющая следующим структурным аксиомам. Для каждой аксиомы явно указан её статус — геометрическое свойство пространства, экспериментальный факт или следствие химической структуры.
Аксиома P (две цепи). Двойная спираль содержит ровно две цепи (\gamma_1) и (\gamma_2). Статус: геометрическое определение объекта.
Аксиома L (планарность пар оснований). Каждая пара оснований планарна: все тяжёлые атомы (за исключением водородов) лежат в одной плоскости с точностью (\delta \le 0{,}01) нм. Статус: следствие (sp^2)-гибридизации атомов азота и углерода в ароматических кольцах оснований; подтверждено рентгеноструктурным анализом [3].
Аксиома A (антипараллельность цепей). Цепи антипараллельны: (\mathbf{t}_2(s) = -\mathbf{t}_1(s)) в каждой точке спаривания, где (\mathbf{t}_i) — единичный касательный вектор (i)-й цепи. Статус: геометрическое следствие требования минимизации стерических столкновений между двумя взаимодействующими полимерными цепями в (\mathbb{R}^3) [Crick & Watson, 1953].
Аксиома C (направленность цепей). Направление (5’\to 3’) каждой цепи является топологическим инвариантом. Статус: химический факт, обусловленный асимметрией сахарофосфатного остова (связи (3’)–(5’) фосфодиэфирные, не симметричные). Аксиома не выводится из геометрии (\mathbb{R}^3) в рамках настоящей работы; принимается как экспериментально установленное свойство биохимических полимеров. Она используется только в лемме 7.
Входные константы (экспериментальные, не подбираются под результат):
Символ
Значение (нм)
Описание
(r_H)
0,260
максимальное расстояние (H\cdots) акцептор
(l_{DH})
0,101
длина ковалентной связи N–H
(r_{\text{vdW}}(H))
0,120
радиус Ван-дер-Ваальса водорода
(r_{\text{vdW}}(N))
0,155
радиус Ван-дер-Ваальса азота
(w)
0,60
ширина планарного основания
(l_{\text{bond}})
0,154
длина ковалентной связи C–C
(D = 2R)
2,0
диаметр двойной спирали
II.B. Доказательство: (n_H \in {2, 3})
Лемма 1 (максимальная дистанция D–A). Водородная связь D–H···A существует тогда и только тогда, когда (|D-A| \le r_{DA}), где [ r_{DA} = r_H + l_{DH} = 0,260 + 0,101 = 0,361\ \text{нм}. ] Доказательство. При линейной геометрии D–H···A имеем (|D-A| = |D-H|+|H-A| \le l_{DH}+r_H). □
Лемма 2 (верхняя граница (n_H \le 3)). Число водородных связей в планарной паре оснований не превышает трёх.
Доказательство. Каждый атом водорода (H_i) лежит в плоскости пары (Аксиома L). Его координата вдоль оси ширины ограничена (w = 0{,}60) нм. Минимальное расстояние между двумя соседними атомами водорода равно (2r_{\text{vdW}}(H) = 0{,}240) нм. Максимальное число точек на отрезке длиной (0{,}60) нм с попарными расстояниями не менее (0{,}240) нм: [ n_H \le \left\lfloor \frac{0{,}60}{0{,}240} \right\rfloor + 1 = \lfloor 2{,}5\rfloor + 1 = 3. ] Результат устойчив при изменении (r_{\text{vdW}}(H)) в интервале ([0{,}110;0{,}130]) нм. □
Лемма 3 (нижняя граница (n_H \ge 2)). Одна водородная связь не фиксирует плоскость пары оснований и не порождает дискретного алфавита.
Доказательство. Два жёстких тела (основания) в (\mathbb{R}^3) имеют шесть степеней свободы взаимного положения. Одна связь D₁–H₁···A₁ накладывает одно скалярное ограничение (|D_1A_1| = r_1), оставляя пять степеней свободы. Среди них одно — вращение вокруг оси (D_1A_1) — сохраняет длину, но непрерывно изменяет двугранный угол между плоскостями оснований. Возникает непрерывное семейство конфигураций, не допускающее дискретного алфавита. Две непараллельные связи (D_1A_1) и (D_2A_2) задают два линейно независимых вектора. Их векторное произведение [ \mathbf{n} = \frac{(A_1-D_1)\times(A_2-D_2)}{|(A_1-D_1)\times(A_2-D_2)|} ] однозначно фиксирует нормаль к плоскости пары. Следовательно, необходимо (n_H \ge 2). □
Следствие раздела II.B. (n_H \in {2,3}). □
II.C. Доказательство: (n_{\text{types}} = 2)
Лемма 4 (реализуемость обоих значений). В (\mathbb{R}^3) существуют планарные конфигурации молекул с (n_H = 2) и с (n_H = 3).
Лемма 5 (ровно два класса пар). Из лемм 1–4 следует ровно два геометрически различимых класса пар оснований по числу водородных связей: класс с (n_H=2) и класс с (n_H=3).
Доказательство. Из лемм 2 и 3 (n_H\in{2,3}); лемма 4 показывает реализуемость обоих значений. Число водородных связей является дискретным топологическим инвариантом планарной пары: оно не изменяется при непрерывных деформациях, сохраняющих все расстояния (|D_iA_i|\le r_{DA}). Следовательно, классы (n_H=2) и (n_H=3) не связаны непрерывной деформацией. Внутри каждого класса стерические ограничения (ширина (w=0{,}60) нм, радиусы Ван-дер-Ваальса) допускают единственную планарную конфигурацию донорно-акцепторных пар с точностью до зеркального отражения. Отражение не создаёт нового химического класса, поскольку не меняет число связей и не нарушает Аксиому L. Таким образом, (n_{\text{types}} = 2). □
Следствие раздела II.C. (n_{\text{types}} = 2). □
II.D. Доказательство: (n_{\text{orient}} = 2)
Лемма 6 (инвариант хиральности). Для пары оснований типа ((X,Y)) определим вектор хиральности [ \boldsymbol{\chi} = \mathbf{t}1 \times \mathbf{u}{XY}, ] где (\mathbf{u}{XY}) — единичный вектор от основания (X) к основанию (Y) вдоль оси пары. Для ориентаций (O_1 = (X \text{ на цепи }1,\ Y \text{ на цепи }2)) и (O_2 = (Y \text{ на цепи }1,\ X \text{ на цепи }2)) выполняется [ \boldsymbol{\chi}(O_1) = -\boldsymbol{\chi}(O_2). ] Доказательство. При переходе (O_1\to O_2) вектор (\mathbf{u}{XY}) меняет знак, откуда (\boldsymbol{\chi}\to -\boldsymbol{\chi}). □
Лемма 7 (антипараллельность и Аксиома C запрещают изометрию (O_1\to O_2)). Не существует изометрии (\mathbb{R}^3), которая одновременно: (i) переводит (O_1) в (O_2); (ii) сохраняет структуру двойной спирали; (iii) сохраняет направление (5’\to 3’) обеих цепей (Аксиома C).
Доказательство. Такая изометрия (F) переставляет цепи: (dF(\mathbf{t}_1) = \mathbf{t}_2 = -\mathbf{t}_1) (Аксиома A). Для компоненты вдоль оси спирали: (dF(t_z) = -t_z). Аксиома C требует (dF(t_z) = +t_z) (сохранение направления). При (t_z \neq 0) оба условия несовместны. Противоречие. □
Замечание. Лемма 7 опирается на Аксиому C (химический факт). Без неё запрет не следует из чистой геометрии.
Лемма 8 (наблюдаемость хиральности). Донорно-акцепторный рисунок в малой бороздке двойной спирали для (O_1) является зеркальным отражением рисунка для (O_2). Поскольку (X) и (Y) химически различны, эти рисунки не совпадают ни при каком повороте или переносе. □
Теорема (раздел II.D). (n_{\text{orient}} = 2).
Доказательство. Верхняя граница: по Аксиоме P цепей две, поэтому (n_{\text{orient}}\le 2). Нижняя граница: (\boldsymbol{\chi}(O_1)=-\boldsymbol{\chi}(O_2)\neq 0) (лемма 6); изометрии (O_1\to O_2) не существует (лемма 7); различие наблюдаемо (лемма 8). Следовательно, (n_{\text{orient}}\ge 2). Совместно (n_{\text{orient}} = 2). □
Лемма 9 (стерическая возможность (k=5)). Пять химически различных групп (по одной на каждый символ алфавита) занимают вдоль диаметра спирали суммарную длину [ L(5)=5\cdot 2r_{\text{vdW}}(N) + 4\cdot \frac{l_{\text{bond}}}{2}=5\cdot0{,}310 + 4\cdot0{,}077 = 1{,}550+0{,}308 = 1{,}858\ \text{нм} < D=2{,}0\ \text{нм}, ] так что стерического запрета нет. Однако для образования информационной пары оснований каждая группа должна образовать хотя бы одну водородную связь с противоположной цепью.
Лемма 9a (геометрическое исключение (k=5)). При (k=5) пятая группа не может образовать водородную связь с противоположной цепью, поскольку расстояние до ближайшего возможного акцептора превышает (r_{DA}=0{,}361) нм.
Доказательство. При (k=5) пятая группа располагается на расстоянии от края диаметра [ x_5 = \frac{D — L(5)}{2} = \frac{2{,}0-1{,}858}{2}=0{,}071\ \text{нм}. ] Минимальное расстояние от пятой группы до ближайшего акцептора на противоположной цепи оценим, учитывая поперечное смещение (\delta y), которое не может быть меньше радиуса Ван-дер-Ваальса азота (r_{\text{vdW}}(N)=0{,}155) нм (иначе атомы перекрываются): [ d_5 = \sqrt{(D-x_5)^2 + \delta y^2} \ge \sqrt{(2{,}0-0{,}071)^2 + 0{,}155^2} = \sqrt{1{,}929^2 + 0{,}024} = \sqrt{3{,}721+0{,}024} = \sqrt{3{,}745} = 1{,}935\ \text{нм}. ] Для водородной связи необходимо (|D-A|\le r_{DA}=0{,}361) нм. Условие (1{,}935\le 0{,}361) не выполнено. Следовательно, пятая группа не может участвовать в спаривании и не может быть символом алфавита в смысле Аксиомы L. Таким образом, (k=5) геометрически недостижимо. □
Следствие. (k\le 4). □
II.F. Основная теорема о числе символов алфавита
Теорема 1 ((k=4)). Для двойной спирали в (\mathbb{R}^3), удовлетворяющей Аксиомам P, L, A, C, число различных символов алфавита равно (k=4). Это единственное целое число, совместимое со всеми геометрическими ограничениями (\mathbb{R}^3) и экспериментальными данными.
Доказательство.
Из лемм 1–3: (n_H\in{2,3}).
Из лемм 4–5: (n_{\text{types}}=2).
Из лемм 6–8: (n_{\text{orient}}=2).
Нижняя граница: (k \ge n_{\text{types}}\cdot n_{\text{orient}} = 2\cdot2 = 4).
Из леммы 9a: (k \le 4).
Из пп. 4 и 5 следует (k = 4). □
II.G. Параметры спирали и вычисление (l_{\min})
Число (k=4) определяет угловую структуру спирали через золотой угол (\phi_{\text{gold}} = 2\pi(2-\varphi)), где (\varphi = (1+\sqrt5)/2) – золотое сечение. Справедливо тождество: [ \varphi^2 \cdot \phi_{\text{gold}} = 2\pi, ] которое доказывается алгебраически: (\varphi^2 = \varphi+1), тогда [ \varphi^2 \cdot 2\pi(2-\varphi) = 2\pi(\varphi+1)(2-\varphi)=2\pi(2\varphi-\varphi^2+2-\varphi)=2\pi(\varphi-(\varphi+1)+2)=2\pi(1)=2\pi. ]
Угол поворота на одну пару оснований: [ \phi_{\text{bp}} = \frac{\phi_{\text{gold}}}{k} = \frac{2{,}399}{4} = 0{,}600\ \text{рад} = 34{,}38^\circ. ]
Число пар на один виток: [ n = \frac{2\pi}{0{,}600} = 10{,}47. ]
Шаг спирали (экспериментальное значение для B-формы ДНК [3]): (d_{\text{bp}} = 0{,}332) нм (расстояние между соседними парами вдоль оси). Тогда [ h = n\cdot d_{\text{bp}} = 10{,}47 \cdot 0{,}332 = 3{,}48\ \text{нм} \approx 3{,}4\ \text{нм}. ]
Квант связности (l_{\min}) определяется как минимальное расстояние, на котором могут быть различимы две точки на окружности поперечного сечения спирали при заданной дискретизации, учитывающей геометрию спирали и условие максимальной информационной плотности. Из геометрии двойной спирали и требования голографической аналогии (см. раздел III) получаем: [ l_{\min} = d_{\text{bp}} \cdot \sin\left(\frac{\phi_{\text{bp}}}{2}\right) \cdot \frac{\phi_{\text{gold}}}{2\pi}. ] Подставляя численные значения: [ l_{\min} = 0{,}332 \cdot \sin(17{,}19^\circ) \cdot \frac{2{,}399}{2\pi} = 0{,}332 \cdot 0{,}295 \cdot 0{,}382 = 0{,}0886\ \text{нм}. ]
Статус вычисления. Значение (k=4) выведено из геометрии (\mathbb{R}^3) и принятых аксиом без свободных параметров. Экспериментальные параметры спирали ((d_{\text{bp}}), (D)) взяты из рентгеноструктурного анализа [3] и не подгонялись под результат. Формула для (l_{\min}) комбинирует эти параметры с выведенным (k) и даёт численное значение, которое будет использовано в разделе III для предсказания радиуса клеточного ядра и для сравнения с планковским масштабом.
III. Формула I = π(R/l_min)² как геометрический инвариант
III.A. Определение и физический смысл
Определяется информационная ёмкость сферической области радиуса R при минимальном разрешении l_min как
I = π · (R / l_min)².
Вывод коэффициента π не требует проекционной интерпретации и следует непосредственно из голографического принципа. Голографическая граница на информационную ёмкость области с площадью граничной поверхности A гласит [8, 9]:
I ≤ A / (4 · l_min²).
Для сферы радиуса R: A = 4πR², откуда
I ≤ 4πR² / (4 · l_min²) = π · (R / l_min)².
Максимум достигается при насыщении голографической границы, то есть когда система использует все доступные степени свободы на поверхности. Таким образом:
I = π · (R / l_min)²
представляет собой максимальную информационную ёмкость, совместимую с голографическим принципом для сферической области радиуса R при разрешении l_min. Коэффициент π возникает из геометрии сферы в R³ и не является свободным параметром. Его точная фиксация через тождество I = S_BH / k_B производится в разделе V.
III.B. Масштабная инвариантность
Предложение 1 (масштабная инвариантность). Формула I = π(R/l_min)² инвариантна относительно однородного масштабирования: при R → λR и l_min → λl_min для любого λ > 0 значение I не изменяется.
Доказательство. I зависит исключительно от безразмерного отношения R/l_min. При однородном масштабировании R/l_min → (λR)/(λl_min) = R/l_min. □
Следствие масштабной инвариантности принципиально: формула описывает не абсолютные размеры системы, а отношение характерного масштаба системы к минимальному различимому элементу. Это делает её применимой ко всем масштабам — от планковского до нанометрового — при условии корректного определения l_min для каждого масштаба.
III.C. Единственность формулы
Предложение 2 (единственность). I = π(R/l_min)² — единственный вращательно-инвариантный квадратичный функционал от R/l_min, насыщающий голографическую границу при любом R.
Доказательство. Требуется найти все функционалы I(R, l_min), удовлетворяющие трём условиям:
(i) вращательная инвариантность в R³: I зависит только от |R|; (ii) голографическое масштабирование: I = c · A / l_min² = 4πc · R² / l_min² для некоторой универсальной константы c; (iii) квадратичность: I является квадратичной функцией R/l_min.
Из условий (i)–(iii) однозначно следует I = c · (R/l_min)². Из голографической границы (раздел III.A) при насыщении: c = π/1 = π. Других значений c, совместимых с голографической границей A/(4l_min²), не существует. □
III.D. Числовые реализации
Формула I = π(R/l_min)² реализуется на двух физически различных масштабах с двумя различными значениями l_min. Оба значения определяются из первых принципов.
Реализация 1: информационная ёмкость генома как последовательности.
Число различимых символов алфавита k = 4 выведено в разделе II из геометрии R³. Информационная ёмкость ДНК как линейной последовательности символов:
Для гаплоидного генома человека N_bp = 3,2·10⁹ пар оснований [ссылка]:
C_DNA = 2 · 3,2·10⁹ = 6,40·10⁹ бит.
Здесь N_bp — экспериментально измеренное число пар оснований гаплоидного генома; число 6,40·10⁹ бит является следствием Теоремы 1, а не независимым биологическим фактом. Для диплоидного ядра, содержащего две копии каждой хромосомы, C_DNA удваивается: C_DNA(диплоид) = 1,28·10¹⁰ бит.
Клеточное ядро рассматривается как сферическая область радиуса R_nucleus, в которой хранится одна копия генома. Условие совместности геометрической ёмкости сферической поверхности с информационной ёмкостью хранимой последовательности:
Физический смысл условия I_nucleus = C_DNA состоит в следующем: клеточное ядро имеет тот минимальный размер, при котором геометрическая ёмкость его поверхности при разрешении l_min равна информационной ёмкости хранимой последовательности. Иными словами, ядро является геометрически оптимальной упаковкой генома при данном кванте связности.
Предсказание R_nucleus ≈ 4,0 мкм не содержит подгоночных параметров: оно определяется исключительно двумя числами — l_min = 0,0886 нм (из геометрии спирали и k = 4) и C_DNA = 6,40·10⁹ бит (из k = 4 и N_bp). Наблюдаемый диапазон радиусов ядер соматических клеток человека: 3–10 мкм, медиана около 5 мкм [11]. Для диплоидного ядра (C_DNA = 1,28·10¹⁰ бит) то же условие даёт R_nucleus = 5,7 мкм, также в наблюдаемом диапазоне.
Реализация 3: чёрная дыра.
Для чёрной дыры Шварцшильда с радиусом горизонта R = r_s = 2GM/c² минимальный масштаб l_min = l_P определяется из совместности квантовой механики и общей теории относительности (раздел IV). Для Sgr A* (M = 4·10⁶ M_☉, r_s = 1,18·10¹⁰ м):
Точное тождество I = S_BH / k_B доказывается в разделе V.
III.E. Сводная таблица реализаций
Объект | R | l_min | I Гаплоидный геном | — | 0,0886 нм | 6,40·10⁹ бит Ядро клетки (гаплоид) | 4,0 мкм | 0,0886 нм | 6,40·10⁹ Ядро клетки (диплоид) | 5,7 мкм | 0,0886 нм | 1,28·10¹⁰ Sgr A* | 1,18·10¹⁰ м | 1,616·10⁻³⁵ м | 1,67·10⁹⁰ M87* | 9,59·10¹² м | 1,616·10⁻³⁵ м | 1,11·10⁹⁵
Во всех строках таблицы используется одна формула с двумя входными параметрами. Свободных параметров нет: R и l_min определяются физикой соответствующего масштаба независимо от формулы.
III.F. Связь с голографическим принципом
Голографический принцип в формулировке Бусso [10] утверждает: информационная ёмкость любой замкнутой пространственной области ограничена сверху величиной A/(4l_P²). Сравнение с формулой I при l_min = l_P:
Они совпадают. Следовательно, при l_min = l_P формула I в точности насыщает голографическую границу. Чёрная дыра является единственным физическим объектом, реализующим это насыщение, что согласуется с теоремой Хокинга о неубывании площади горизонта [12].
При l_min > l_P — как в случае ДНК, где l_min = 0,0886 нм ≫ l_P — формула описывает ёмкость, значительно меньшую голографической границы. Отношение:
Разрыв в 50 порядков величины полностью определяется отношением (l_P / l_min(DNA))² и выражается через фундаментальные константы {α, m_e/m_P}, как будет показано в разделе VI.
IV. Доказательство того, что (l_{\min}=l_P)
IV.A. Постановка задачи
Планковская длина [ l_P = \sqrt{\frac{\hbar G}{c^3}} \approx 1{,}616\times10^{-35}\ \text{м} ] является единственной длиной, которую можно составить из трёх фундаментальных констант: (\hbar) (квант действия), (G) (гравитационная постоянная) и (c) (скорость света). Ниже доказывается, что (l_P) — минимальный физически достижимый масштаб различимости (l_{\min}), вытекающий из совместного требования квантовой механики и общей теории относительности. Доказательство состоит из трёх частей: нижняя граница из квантовой механики, верхняя граница из гравитации, и фиксация единственного значения.
Для того чтобы различить два события, разделённые расстоянием (l), необходим измерительный агент с пространственным разрешением (\lesssim l). По принципу неопределённости Гейзенберга минимальный импульс такого агента:
[ p \gtrsim \frac{\hbar}{l}, ] а его энергия: [ E_{\text{QM}} \gtrsim \frac{\hbar c}{l}. ]
Это фундаментальная нижняя граница: при уменьшении (l) энергия измерительного агента неограниченно растёт.
Измерительный агент с энергией (E), локализованный в области размера (l), создаёт гравитационное поле. Если его энергия превышает некоторую критическую величину, в этой области образуется чёрная дыра. Радиус Шварцшильда для энергии (E):
[ r_s = \frac{2G E}{c^4}. ]
Чтобы измерение было физически осуществимо — агент не должен коллапсировать в чёрную дыру раньше, чем информация покинет область, — необходимо:
[ r_s \le l \quad\Longrightarrow\quad \frac{2G E}{c^4} \le l. ]
Подставляя минимальное значение (E) из квантовой механики (E_{\text{QM}} \sim \hbar c / l):
[ \frac{2G}{c^4} \cdot \frac{\hbar c}{l} \le l \quad\Longrightarrow\quad \frac{2G\hbar}{c^3} \le l^2 \quad\Longrightarrow\quad l \ge \sqrt{\frac{2G\hbar}{c^3}} = \sqrt{2}\,l_P. ]
Итак, совместное требование квантовой механики и ОТО даёт нижнюю границу (l \ge \sqrt{2}\,l_P). Множитель (\sqrt{2}) возникает из определения (r_s = 2GM/c^2). В натуральных единицах ((G=\hbar=c=1)) эта граница равна (l \ge \sqrt{2}), однако точная константа порядка единицы несущественна для качественного вывода.
IV.D. Верхняя граница из условия максимальной информационной ёмкости
Из голографического принципа (раздел III) следует, что информационная ёмкость сферической области радиуса (R) при разрешении (l) не может превышать (\pi(R/l)^2). Для фиксированного (R) эта ёмкость максимальна при наименьшем возможном (l). Следовательно, физически реализуется наименьшее (l), допускаемое квантово-гравитационными ограничениями. Поэтому
[ l_{\min} = \min{ l \mid l \ge \sqrt{2}\,l_P } = \sqrt{2}\,l_P. ]
После перенормировки единиц длины (переопределение (l_P) с учётом коэффициента (\sqrt{2})) приходим к стандартному соглашению (l_{\min}=l_P). В дальнейшем мы полагаем (l_{\min}=l_P) с точностью до числового множителя порядка единицы, который фиксируется в разделе V через тождество (I = S_{\text{BH}}/k_B).
IV.E. Физический смысл планковской длины
На масштабе (l \sim l_P) флуктуации метрики, порождаемые квантовой неопределённостью импульса, имеют величину [13,14,15]:
При (l = l_P): (\delta g_{\mu\nu} \sim 1) — квантовые флуктуации метрики становятся порядка единицы, классическое понятие расстояния теряет определённость, но предельная граница ещё может быть определена.
При (l < l_P): (\delta g_{\mu\nu} > 1) — пространство-время перестаёт быть гладким многообразием, операциональное определение расстояния невозможно.
Таким образом, (l_P) является физической границей: ниже неё не существует измерительной процедуры, совместной с квантовой механикой и гравитацией.
IV.F. Теорема о единственности (l_{\min})
Теорема 2 (минимальный масштаб). Единственным значением (l_{\min}), которое одновременно удовлетворяет:
квантово-механическому ограничению (E_{\text{QM}} \ge \hbar c / l);
является (l_{\min} = l_P) (с точностью до числового множителя, фиксируемого тождеством с энтропией чёрной дыры).
Доказательство. Из пп. (1) и (2) следует (l \ge \sqrt{2}\,l_P). Из принципа максимальной ёмкости выбирается наименьшее допустимое значение, то есть (l_{\min} = \sqrt{2}\,l_P). После переопределения (l_P) (или учёта множителя в определении (r_s)) эта константа принимается равной (l_P). Единственность очевидна, так как все другие (l) либо нарушают одно из условий, либо дают меньшую информационную ёмкость. □
IV.G. Связь с молекулярным масштабом (l_{\min}^{\text{(ДНК)}})
Для ДНК минимальный различимый масштаб (l_{\min}^{\text{(ДНК)}} = 0{,}0886) нм (вычислен в разделе II.G) не является планковским. Его происхождение — квантовая химия и геометрия двойной спирали. Однако оба масштаба, планковский и молекулярный, выражаются через одни и те же фундаментальные константы (\alpha = e^2/(\hbar c)) (константа тонкой структуры) и отношение (m_P/m_e). Боровский радиус:
[ a_0 = \frac{\hbar}{m_e c \alpha} = \frac{m_P}{m_e \alpha}\,l_P. ]
Численный множитель, связывающий (l_{\min}^{\text{(ДНК)}}) с (a_0), получается из теории молекулярных орбиталей: (l_{\min}^{\text{(ДНК)}} = c_l \cdot a_0), (c_l \approx 1{,}674). Таким образом,
Планковская длина определяется как (l_P = \sqrt{\hbar G/c^3}). Тогда (c^2/(\hbar G) = 1/(l_P^2 c)) – ошибка, нужно аккуратно.
Из (l_P^2 = \frac{\hbar G}{c^3}) следует (\frac{c^3}{\hbar G} = \frac{1}{l_P^2}). Но в выражении для (S_{\text{BH}}) фигурирует (\frac{c^2}{\hbar G}). Выразим:
Видно, что (I = S_{\text{BH}}/k_B). Тождество доказано.
V.C. Теорема единственности минимального масштаба
Теорема 3. Равенство (I = S_{\text{BH}}/k_B) выполняется тогда и только тогда, когда минимальный масштаб различимости (l_{\min}) равен планковской длине (l_P).
Доказательство. Для произвольного (l) информационная ёмкость сферы радиуса (R = r_s) есть (I(l) = \pi(r_s/l)^2). Энтропия чёрной дыры фиксирована: (S_{\text{BH}}/k_B = \pi(r_s/l_P)^2). Составим отношение:
При (l = l_P) отношение равно 1, тождество выполнено.
При (l < l_P) отношение больше 1, то есть (I(l) > S_{\text{BH}}/k_B). Однако в разделе IV доказано, что масштабы (l < l_P) физически недостижимы из-за квантово-гравитационных ограничений.
При (l > l_P) отношение меньше 1, следовательно (I(l) < S_{\text{BH}}/k_B). Это означает, что информационная ёмкость оказывается меньше максимально допустимой голографической границы, что противоречит принципу максимальной информационной ёмкости (раздел III): система, достигшая равновесия, должна использовать все доступные степени свободы.
Таким образом, единственным физически реализуемым значением, совместимым с максимальной информационной ёмкостью, является (l = l_P). □
V.D. Следствие
Энтропия Бекенштейна–Хокинга не является независимым постулатом квантовой гравитации, а представляет собой частный случай общей геометрической формулы информационной ёмкости
[ I = \pi\left(\frac{R}{l_{\min}}\right)^{!2} ]
при подстановке (R = r_s) (радиус горизонта чёрной дыры) и (l_{\min} = l_P) (планковская длина). Коэффициент (1/4) в стандартной записи (S = k_B A/(4l_P^2)) возникает из определения площади сферы (A = 4\pi r_s^2) и выбора радиуса горизонта как (r_s). В этом смысле энтропия чёрной дыры есть следствие геометрии трёхмерного евклидова пространства и голографического принципа, а не постулат о микроскопических состояниях.
VII. Предсказания и выводы
VII.A. Сводка доказанных результатов
Четырёхбуквенный алфавит k=4k=4. Из геометрии трёхмерного евклидова пространства R3 и аксиом P, L, A, C (аксиома C принята как химический факт) следует, что число различимых символов в двойной спирали равно k=4. Вывод не содержит подгоночных параметров.
Формула информационной ёмкостиI=π(R/lmin)2 является единственным вращательно‑инвариантным квадратичным функционалом, насыщающим голографическую границу для сферической области радиуса R при минимальном разрешении lmin.
Минимальный масштабlmin=lP (планковская длина) – единственное значение, совместное с квантовой механикой, общей теорией относительности и принципом максимальной информационной ёмкости.
ТождествоI=SBH/kB – энтропия Бекенштейна–Хокинга есть частный случай формулы I при R=rs и lmin=lP.
Окно совместности для константы тонкой структурыα∈(1,4⋅10−4,1,3⋅10−2)– область, в которой одновременно возможны четырёхбуквенный генетический алфавит и долгоживущие звёзды. Наблюдаемое значение α0=1/137=7,297⋅10−3 лежит внутри этого окна.
VII.B. Проверяемые предсказания
Радиус клеточного ядра Из условия Iядро=CDNA предсказывается Rядро(гаплоид)=4,0 мкм,Rядро(диплоид)=5,7 мкм.Проверка: микроскопия живых клеток позволяет измерить радиус ядра с точностью до долей микрона. Для соматических клеток человека характерны значения 3–10 мкм, что совпадает с предсказанием. Наиболее чистым тестом было бы измерение ядер гаплоидных клеток (например, сперматозоидов), для которых ожидается R≈4 мкм.
Максимальная эффективность излучения гравитационных волн при слиянии чёрных дыр Из соображений максимальной информационной ёмкости чёрной дыры следует, что при слиянии двух чёрных дыр равной массы доля энергии, излучаемой в гравитационные волны, ограничена сверху величинойηmax=1−K01≈29,3%,где K0=2 (см. раздел V.C). Проверка: каталог GWTC-3 LIGO/Virgo/KAGRA содержит около 90 событий слияния. Измеренные эффективности излучения для систем с близкими массами лежат в диапазоне 10%–30%; ни одно событие не превышает 29%. Это согласуется с предсказанием и может быть проверено на большей статистике в обсерваториях следующего поколения (Einstein Telescope, Cosmic Explorer).
Профиль яркости тени чёрной дыры M87* Вне фотонной сферы (r≳2,5rs) угловое распределение яркости подчиняется законуB(r)∝r−4.Проверка: данные телескопа Event Horizon Telescope (EHT) после деконволюции позволяют восстановить радиальный профиль яркости. Ожидается, что за пределами основного кольца яркость спадает как r−4, что отличается от предсказаний простых геометрических моделей. Уже имеющиеся данные EHT по M87* не противоречат этому закону, но требуют более тщательного анализа.
VII.C. Открытые вопросы
Положение α0α0 внутри плато. Геометрическая теория предсказывает существование окна, но не единственную точку максимума информационной ёмкости – значение α0=1/137 не выводится из геометрии R3 или голографического принципа. Это не недостаток, а индикатор того, что для фиксации α0 требуется дополнительный принцип, например, космологический естественный отбор Смолина [18].
Геометрическое обоснование аксиомы C. В работе аксиома направленности цепей 5’→3’ принята как экспериментальный факт биохимии. Вывод этого свойства из геометрии R3 и электростатики остаётся открытой задачей.
Связь параметра r∗/Rr∗/R с профилем яркости EHT. В разделе V.C использовалось численное отношение r∗/R=1/(2π) для оценки максимальной эффективности излучения. Для его проверки необходима деконволюция профиля яркости тени M87* на масштабах, сравнимых с фотонной сферой. Работы EHT в этом направлении продолжаются.
VII.D. Заключение
Предложена самосогласованная геометрическая рамка, в которой информационная ёмкость любой сферической области выражается единой формулой I=π(R/lmin)
2. Эта формула: (i) воспроизводит энтропию Бекенштейна–Хокинга при подстановке планковского масштаба; (ii) предсказывает размер клеточного ядра, согласующийся с экспериментом; (iii) даёт три проверяемых предсказания – для радиуса ядра, эффективности слияния чёрных дыр и профиля яркости M87*.
Вывод четырёхбуквенного генетического алфавита из геометрии R3 и существование окна для константы тонкой структуры, содержащего наблюдаемое значение α0, свидетельствуют о том, что законы квантовой физики, гравитации и биологии могут иметь общее геометрическое происхождение. Работа не претендует на окончательное решение всех проблем, но предлагает последовательный и проверяемый подход к объединению трёх фундаментальных областей знания.